Articles
- Page Path
- HOME > Endocrinol Metab > Volume 29(2); 2014 > Article
-
Original ArticleA Novel Cytosolic Isoform of Mitochondrial Trans-2-Enoyl-CoA Reductase Enhances Peroxisome Proliferator-Activated Receptor α Activity
- Dong-Gyu Kim1, Jae Cheal Yoo1, Eunju Kim2, Young-Sun Lee1, Oleg V. Yarishkin2, Da Yong Lee2, Kun Ho Lee3, Seong-Geun Hong1, Eun Mi Hwang2, Jae-Yong Park1,2
-
Endocrinology and Metabolism 2014;29(2):185-194.
DOI: https://doi.org/10.3803/EnM.2014.29.2.185
Published online: June 26, 2014
1Department of Physiology, Institute of Health Science, Medical Research Center for Neural Dysfunction, Gyeongsang National University School of Medicine, Jinju, Korea.
2Center for Functional Connectomics, Korea Institute of Science and Technology, Seoul, Korea.
3Department of Marine Life Science, Chosun University College of Natural Sciences, Gwangju, Korea.
- Corresponding author: Jae-Yong Park. Department of Physiology, Gyeongsang National University School of Medicine, 15 Jinju-daero 816beon-gil, Jinju 660-751, Korea. Tel: +82-55-772-8043, Fax: +82-55-772-8049, jaeyong@gnu.ac.kr
• Received: August 13, 2013 • Accepted: October 11, 2013
Copyright © 2014 Korean Endocrine Society
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
ABSTRACT
-
Background
- Mitochondrial trans-2-enoyl-CoA reductase (MECR) is involved in mitochondrial synthesis of fatty acids and is highly expressed in mitochondria. MECR is also known as nuclear receptor binding factor-1, which was originally reported with yeast two-hybrid screening as a binding protein of the nuclear hormone receptor peroxisome proliferator-activated receptor α (PPARα). However, MECR and PPARα are localized at different compartment, mitochondria, and the nucleus, respectively. Therefore, the presence of a cytosolic or nuclear isoform of MECR is necessary for functional interaction between MECR and PPARα.
-
Methods
- To identify the expression pattern of MECR and the cytosolic form of MECR (cMECR), we performed reverse transcription polymerase chain reaction (RT-PCR) with various tissue samples from Sprague-Dawley rats. To confirm the interaction between cMECR and PPARα, we performed several binding assays such as yeast two-hybrid, coimmunoprecipitation, and bimolecular fluorescence complementation. To observe subcellular localization of these proteins, immunocytochemistry was performed. A luciferase assay was used to measure PPARα activity.
-
Results
- We provide evidence of an alternatively spliced variant of the rat MECR gene that yields cMECR. The cMECR lacks the N-terminal 76 amino acids of MECR and shows uniform distribution in the cytoplasm and nucleus of HeLa cells. cMECR directly bound PPARα in the nucleus and increased PPARα-dependent luciferase activity in HeLa cells.
-
Conclusion
- We found the cytosolic form of MECR (cMECR) was expressed in the cytosolic and/or nuclear region, directly binds with PPARα, and enhances PPARα activity.
- Mammalian mitochondria perform nicotinamide adenine dinucleotide phosphate (NADPH)-dependent de novo fatty acid synthesis (FAS) [1]. This mitochondrial FAS pathway resembles the well-understood bacterial FAS II pathway [2]. Mitochondrial trans-2-enoyl-CoA reductase (MECR) is a component of the mitochondrial FAS II pathway and catalyzes the fatty acid elongation cycle in the last step, which is the NADPH-dependent reduction of the enoyl-acyl carrier protein substrate [3,4,5]. MECR is primarily localized in mitochondria [6,7,8], and the N-terminal amino acids of this protein are important for its mitochondrial localization [9]. MECR is also known as nuclear receptor binding factor-1 (NRBF-1) because, using yeast two-hybrid screening, it was identified as a binding protein of the nuclear hormone receptor peroxisome proliferator-activated receptor α (PPARα) [10]. However, the biological meaning of the interaction between MECR and PPARα has not been determined.
- PPARα is a ligand-activated transcription factor that is one of three different PPAR subtypes: PPARα, PPARβ/δ, and PPARγ. The PPARs play important roles in nutrient homeostasis [11,12,13] and are localized in the nucleus. Although MECR was previously reported as a binding protein of PPARα [10], interaction between MECR and PPARα seems not to occur in mammalian cells due to their different subcellular localizations, mitochondria and the nucleus, respectively. Therefore, the presence of a cytosolic or nuclear isoform of MECR is necessary for functional interaction between MECR and PPARα in the nuclei of cells.
- Here, we analyzed the expression pattern of MECR in several rat tissues and found a novel splice variant of MECR in which an additional exon was inserted between exon 1 and exon 2. The protein generated from this splicing variant has an N-terminal region that does not contain the mitochondrial targeting signal peptide and thus is not localized in mitochondria. Moreover, this MECR variant bound PPARα in the nucleus and enhanced PPARα transcriptional activity. Based on these results, we propose that this novel variant of MECR, cytosolic MECR (cMECR), plays a role in intracellular signal pathways as an interacting partner of PPARα.
INTRODUCTION
- RNA extraction and reverse transcription polymerase chain reaction
- Total RNA was extracted from tissues of male Sprague-Dawley rats (14 weeks old) using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's protocol. Total RNA (1 µg) from each sample was reverse transcribed using random primers (50 pmol), SuperScript III Reverse Transcriptase (Invitrogen), and dNTPs (1 mM) at 42℃ for 1 hour. The forward primer (5'-ATGTTGGTCAGCCGGCGACT-3') and reverse primer (5'-TCACATAGTGAGAATCTGCT-3') were designed to amplify full-length MECR. Additional primers were used for specific detection of MECR and cMECR cDNA: forward primer (5'-GTGCTGGAAGCGGCATGTTG-3') and reverse primer (5'-TGAGCTCCAGGTTCTTCAGT-3'). cMECR and PPARα fragments were amplified under the following cycle conditions: denaturation at 94℃ for 30 seconds, annealing at 55℃ for 30 seconds, and extension at 72℃ for 30 seconds. This cycle was repeated 35 times. Glyceraldehyde 3-phosphate dehydrogenase fragments were also amplified under the same conditions except that 25 cycles were run. Polymerase chain reaction (PCR) products were analyzed with 2.0% agarose gel electrophoresis, purified, and ligated into the pGEM-T Easy vector (Promega, Madison, WI, USA). The recombinant plasmids were sequenced.
- Construction of plasmids and antibodies
- The rat full-length MECR (GenBank accession no. AB015724), cytosolic variant of MECR, and PPARα (GenBank accession no. NM_001001928) were cloned using the Gateway Cloning System (Invitrogen) as previously described [14]. After amplification of the three genes, PCR products were cloned into the pDONR207 vector and then subcloned into self-constructed destination vectors such as pDS_XB-HA, pDs_XB-Flag, pDS_XB-enhanced green fluorescent protein (EGFP), and pDS_XB-mCherry expression vectors. Anti-Flag (F6531) antibody was purchased from Sigma (St. Louis, MO, USA). Anti-EGFP (B-2) and antihemagglutinin (anti-HA) (F-7) antibodies were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
- Cell culture and transfection
- HeLa cells were maintained in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum (Invitrogen) and 1% penicillin-streptomycin in a humid atmosphere containing 5% CO2 at 37℃. When needed, cells were seeded onto cover slips for imaging analysis or 60-mm dishes for preparation of lysates for Western blot analysis. Transfection of expression vectors was performed with lipofectamine 2000 reagent (Invitrogen) according to the manufacturer's protocol. Transfected cells were cultured for an additional 24 hours in growth medium and then used for further analysis.
- Coimmunoprecipitation and Western blot analysis
- Transfected cells were lysed with radioimmunoprecipitation assay buffer (50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 5 mM ethylenediaminetetraacetic acid, 1 mM phenylmethanesulfonyl fluoride, and 1% NP-40) containing a protease inhibitor cocktail (Sigma). Whole-cell lysates were incubated on ice for 30 minutes and then cleared at 13,000 rpm for 20 minutes at 4℃. The immune complexes were incubated for 1 hour at 4℃ with gentle rotation with 20 µL protein A/G PLUS-agarose beads (Santa Cruz Biotechnology) that had been prewashed and suspended in 100 mL cold lysis buffer. The samples were washed three times in 1 mL cold lysis buffer and eluted in 20 µL sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) sample buffer. The proteins were separated with 10% SDS-PAGE and blotted onto polyvinylidene fluoride membranes. The blots were blocked with 5% skim milk in Tris-buffered saline with Tween 20 (20 mM Tris-buffered saline and 0.05% Tween 20, pH 7.5) at room temperature for 20 minutes and incubated overnight at 4℃ with anti-GFP antibody (1:1,000), anti-HA antibody (1:1,000), or anti-FLAG antibody (1:1,000). Blots were then washed and incubated with horseradish peroxidase-conjugated secondary antibody (1: 3,000), followed by washing and detection of immunoreactivity with enhanced chemiluminescence (Amersham, Piscataway, NJ, USA).
- Imaging analysis
- Subcellular distribution of MECR, cMECR, and PPARα were confirmed according to the EGFP and mCherry fluorescence detection method with a confocal microscope (Olympus Fluoview FV1000, Olympus, Tokyo, Japan). For imaging analysis, cells were plated onto glass cover slips. After transfection, the cells were grown for 24 hours and treated with the mitochondrion-selective fluorescent dye mito-tracker red CMXRos (Molecular Probes Europe BV, Leiden, Netherlands) according to the manufacturer's instructions.
- Yeast two-hybrid assay
- cMECR was cloned into pGADT7 encoding the activation domain, and PPARα was ligated into pGBKT7 encoding the GAL4 DNA binding domain. To evaluate the protein-protein interaction between cMECR and PPARα, both pGAD-cMECR and pGBK-PPARα were cotransformed into the yeast strain AH109. This strain is unable to synthesize histidine. However, interaction between cMECR and PPARα enables the yeast to make the His3 enzyme, thereby permitting histidine biosynthesis and growth on His minimal medium.
- Bimolecular fluorescence complementation assays
- C- and N-terminal venus plasmids were purchased from Addgene and modified to contain cMECR or PPARα with subcloning. Cells were transfected with the lipofectamine 2000 reagent (Invitrogen) with venus plasmids containing cMECR and PPARα. After 24 hours, cells were imaged with confocal microscopy as described above.
- Luciferase assay
- HeLa cells were plated at a density of 1.0 to 1.5×105 cells per well onto six-well dishes. Twenty-four hours later, cells were cotransfected with 1 µg of PPAR responsive element (PPRE)-3x-TK-Luc, the firefly luciferase reporter gene containing PPRE; 0.2 µg of the PPARα expression vector; and either the MECR expression vector or cMECR expression vector. Transfections were accomplished with lipofectamine 2,000 reagent according to the manufacturer's recommendations. The PPRE-3x-TK-Luc plasmid includes three PPREs upstream of an inducible thymidine kinase promoter controlling transcription of the firefly luciferase gene. Cells were harvested 24 hours after the start of exposure to vehicle or inducing agent. Luciferase assays were performed using the Luciferase Assay System (Promega) according to the manufacturer's protocol.
METHODS
- Identification of a novel alternative transcript variant of MECR
- During reverse transcription (RT)-PCR analysis of MECR expression in the rat brain, two different MECR transcripts were detected. One was the expected MECR mRNA (210 bp), which corresponds to the known MECR mRNA (GenBank accession no. AB015724). The other was a longer 306-bp PCR fragment (Fig. 1C). Because PCR was carried out with specific primers corresponding to exons 1 and 2 of rat MECR cDNA, alternative splicing was proposed to account for the presence of this fragment. Fig. 1A shows the 10 exons of rat MECR cDNA. The novel MECR variant has an additional 96 nucleotides between exon 1 and exon 2. Basic local alignment search tool analysis of the aligned sequences of rat genomic DNA (chromosome 5 genomic contig NW_047784) and cDNA of this MECR variant showed that the flanking nucleotides of convergent points between exon 1 and exon 2 followed the GT/AG rule of alternative splicing (Supplementary Fig. 1A). This addition of 96 nucleotides generated a frame-shifted MECR, and the N-terminal 76 amino acids were deleted by use of another start codon in exon 3 (Fig. 1B). Because this splice variant lacked the N-terminal mitochondrial targeting sequences, we named it cMECR.
- Next, to compare expression of cMECR and MECR, RT-PCR analyses were done using RNA samples from eight different tissues with a specific primer set as shown in Fig. 1B. As shown in Fig. 1C, cMECR (306 bp) and MECR (210 bp) were ubiquitously expressed in most tissues examined, although the expression level of cMECR was lower than that of MECR. Furthermore, we found amino acid sequences of human MECR isoform 2 (GenBank accession no. NP001019903.1) were well matched with that of rat cMECR (Supplementary Fig. 1B). These data raise the possibility that cMECR can be produced in other species.
- cMECR localized in the cytoplasm and nucleus of HeLa cells
- To visualize the intracellular distribution of MECR in HEK cells, we generated two constructs containing EGFP-tagged full-length rat MECR. As shown in Supplementary Fig. 2, when EGFP was tagged to the C-terminal end of MECR (MECR-EGFP), this protein was distinctly expressed in mitochondria, as previously reported [3]. However, the N-terminal EGFP-tagged MECR (EGFP-MECR) was diffusely expressed in the cytoplasm of HEK cells. N-terminal tagging of the protein with EGFP might have disrupted the proper mitochondrial localization of MECR, perhaps because the N-terminal mitochondrial targeting sequence of MECR was masked by the EGFP. These results suggest that N-terminal mitochondrial targeting sequences are critical for mitochondrial localization of MECR proteins.
- Because cMECR lacks the N-terminal mitochondrial targeting sequences, the intracellular distribution of this protein was unclear. The intracellular distribution of MECR and cMECR was thus compared by tagging both cDNAs with EGFP at their C-terminal ends and then transiently expressing them in HeLa cells (Fig. 2). MECR-EGFP showed a well-defined punctate pattern that clearly colocalized with mito tracker, a marker of mitochondria (Fig. 2B). However, cMECR-EGFP was expressed in cytosolic and nuclear regions and did not colocalize with Mito tracker (Fig. 2C).
- cMECR directly interacts with PPARα
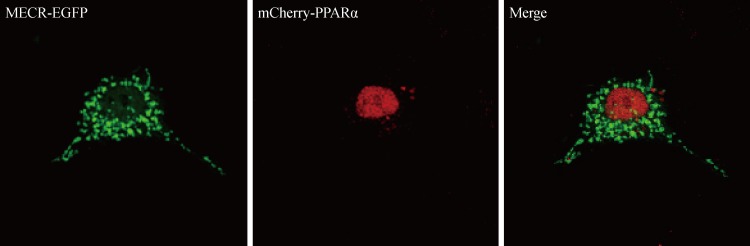
- MECR, also known as NRBF-1, was previously identified with yeast two-hybrid screening as a PPARα-interacting protein [10], although explaining how mitochondrial-targeted MECR interacted with the nuclear hormone receptor PPARα was difficult. In contrast to MECR, cMECR was localized in the cytoplasm and nucleus of HeLa cells, and its amino acid sequence is exactly the same as that of MECR, except for the N-terminal 76 amino acids that compose the mitochondrial targeting sequence. Thus, cMECR, rather than mitochondrial MECR, could interact with PPARα in the nucleus. Indeed, the yeast two-hybrid assay showed a direct interaction between cMECR and PPARα (Fig. 3A). Immunoprecipitation and colocalization analyses also showed that cMECR interacted with PPARα in HeLa cells (Fig. 3B). Furthermore, cMECR-EGFP colocalized with mCherry-tagged PPARα in the nucleus of HeLa cells (Fig. 3C). In contrast, wild-type MECR was not colocalized with PPARα in HeLa cells (Supplementary Fig. 3). Taken together, these data strongly indicate that cMECR, but not MECR, is a bona fide binding partner of PPARα.
- cMECR binds with PPARα in the nucleus and potentiates PPARα activity
- To more clearly investigate whether the interaction between cMECR and PPARα can occur in the nucleus of HeLa cells, we performed a bimolecular fluorescence complementation (BiFC) experiment. The BiFC assay provides a direct approach for the visualization of molecular interactions in living cells [15,16]. We constructed the N-terminal part of venus plasmids tagged to cMECR (cMECR-VN) and the C-terminal portion of venus plasmids tagged to PPARα (VC-PPARα) (Fig. 4A). Consistent with the immunoprecipitation and colocalization analyses, the BiFC experiment also indicated protein-protein interaction between cMECR and PPARα (Fig. 4B). Only when both constructs were cotransfected was the BiFC signal seen in the nucleus of HeLa cells. Individual constructs (cMECR-VN or VC-PPARα) did not produce fluorescence alone (Fig. 4B). These results imply that cMECR interacts with PPARα in the nucleus of HeLa cells.
- cMECR interacts with PPARα, a well-known nuclear hormone receptor. Thus, to examine whether cMECR can modulate PPARα activity, we transfected cMECR into HeLa cells together with PPARα and a luciferase reporter plasmid containing a PPAR-binding site (Fig. 4C). Compared with cells transfected with PPARα only, PPAR-dependent luciferase expression levels were significantly increased in the presence of cMECR. These results indicate that cMECR can act as a coactivator of PPARα in the nucleus of HeLa cells.
RESULTS
- MECR has been reported to be a mitochondrial protein [7] that catalyzes NADPH-dependent reduction of trans-2-enoyl thioesters to the corresponding saturated acyl thioesters [5]. However, NRBF-1 (also known as MECR) was originally identified as a binding protein of the nuclear receptor PPARα [10]. In that paper, the authors did not perform any assessment of endogenous binding in cells or report the biological meaning of this protein-protein interaction, although NRBF-1 binds with PPARα in the yeast two-hybrid system.
- Here, we report a cMECR that is generated by alternative splicing. cMECR is an N-terminal-76-amino-acid truncated isoform of MECR. Because mitochondrial targeting signal sequences are contained between amino acids 1 and 53 of MECR [8], cMECR lacks this mitochondrial targeting signal sequence and was preferentially localizes to cytosolic and nuclear regions and was not present in mitochondria (Fig. 2C). In addition, our data show that N-terminal EGFP-tagged MECR also lost its proper intracellular localization (Supplementary Fig. 2). Therefore, the N-terminal region of MECR is critical for mitochondrial localization of the MECR protein.
- Different intracellular distribution of cMECR strongly implies that its biological functions differ from the functions of MECR. In addition, cMECR mRNA was ubiquitously expressed in eight tested tissues (Fig. 1C), implying that cMECR could perform common essential functions in the various tissues. Although further studies should be performed to elucidate the function(s) of cMECR, we present its first function in this study. Various nuclear receptors activate transcription of their target genes through interactions with coactivators or corepressors [17,18]. Like other coactivators, cMECR directly bound PPARα in the nucleus and potentiated PPARβ activity (Fig. 4).
- Generally, PPARα is found in tissues where fatty acid metabolism is important and regulates genes involved in lipid and lipoprotein metabolism as well as glucose homeostasis [19,20,21]. Because cMECR potentiates PPARα activity in HeLa cells, cMECR could be involved in these biological phenomena. An earlier study used the yeast two-hybrid assay and demonstrated that MECR (NRBF-1) also interacts with various nuclear hormone receptors such as TRβ, RARα, RXRα, and HNF-4 [10]. Because our data strongly support the idea that cMECR is a bona fide binding partner of PPARα and its amino acid sequence is the same as that of MECR except that it lacks the N-terminal 76 amino acids, cMECR might also bind with other nuclear hormone receptors. This possibility should be tested in future studies.
- In summary, our results suggest that cMECR, a novel alternatively spliced variant of MECR, directly interacts with PPARα in the nucleus and could be a positive regulator in the physiological regulation of PPARα.
DISCUSSION
-
Acknowledgements
- This research was supported by the Basic Research Promotion Fund (NRF-2008-313-C00758).
ACKNOWLEDGMENTS
- 1. Chen Z, Leskinen H, Liimatta E, Sormunen RT, Miinalainen IJ, Hassinen IE, Hiltunen JK. Myocardial overexpression of Mecr, a gene of mitochondrial FAS II leads to cardiac dysfunction in mouse. PLoS One 2009;4:e5589ArticlePubMedPMC
- 2. White SW, Zheng J, Zhang YM, Rock . The structural biology of type II fatty acid biosynthesis. Annu Rev Biochem 2005;74:791–831. ArticlePubMed
- 3. Torkko JM, Koivuranta KT, Miinalainen IJ, Yagi AI, Schmitz W, Kastaniotis AJ, Airenne TT, Gurvitz A, Hiltunen KJ. Candida tropicalis Etr1p and Saccharomyces cerevisiae Ybr026p (Mrf1'p), 2-enoyl thioester reductases essential for mitochondrial respiratory competence. Mol Cell Biol 2001;21:6243–6253. ArticlePubMedPMC
- 4. Chen ZJ, Pudas R, Sharma S, Smart OS, Juffer AH, Hiltunen JK, Wierenga RK, Haapalainen AM. Structural enzymological studies of 2-enoyl thioester reductase of the human mitochondrial FAS II pathway: new insights into its substrate recognition properties. J Mol Biol 2008;379:830–844. ArticlePubMed
- 5. Hoffmeister M, Piotrowski M, Nowitzki U, Martin W. Mitochondrial trans-2-enoyl-CoA reductase of wax ester fermentation from Euglena gracilis defines a new family of enzymes involved in lipid synthesis. J Biol Chem 2005;280:4329–4338. ArticlePubMed
- 6. Pfanner N. Protein sorting: recognizing mitochondrial presequences. Curr Biol 2000;10:R412–R415. ArticlePubMed
- 7. Emanuelsson O, Nielsen H, Brunak S, von Heijne G. Predicting subcellular localization of proteins based on their N-terminal amino acid sequence. J Mol Biol 2000;300:1005–1016. ArticlePubMed
- 8. Claros MG, Vincens P. Computational method to predict mitochondrially imported proteins and their targeting sequences. Eur J Biochem 1996;241:779–786. ArticlePubMed
- 9. Chen JQ, Delannoy M, Cooke C, Yager JD. Mitochondrial localization of ERalpha and ERbeta in human MCF7 cells. Am J Physiol Endocrinol Metab 2004;286:E1011–E1022. ArticlePubMed
- 10. Masuda N, Yasumo H, Furusawa T, Tsukamoto T, Sadano H, Osumi T. Nuclear receptor binding factor-1 (NRBF-1), a protein interacting with a wide spectrum of nuclear hormone receptors. Gene 1998;221:225–233. ArticlePubMed
- 11. Desvergne B, Wahli W. Peroxisome proliferator-activated receptors: nuclear control of metabolism. Endocr Rev 1999;20:649–688. ArticlePubMed
- 12. Kersten S, Desvergne B, Wahli W. Roles of PPARs in health and disease. Nature 2000;405:421–424. ArticlePubMedPDF
- 13. Evans RM, Barish GD, Wang YX. PPARs and the complex journey to obesity. Nat Med 2004;10:355–361. ArticlePubMedPDF
- 14. Park JY, Hwang EM, Yarishkin O, Seo JH, Kim E, Yoo J, Yi GS, Kim DG, Park N, Ha CM, La JH, Kang D, Han J, Oh U, Hong SG. TRPM4b channel suppresses store-operated Ca2+ entry by a novel protein-protein interaction with the TRPC3 channel. Biochem Biophys Res Commun 2008;368:677–683. ArticlePubMed
- 15. Hu CD, Chinenov Y, Kerppola TK. Visualization of interactions among bZIP and Rel family proteins in living cells using bimolecular fluorescence complementation. Mol Cell 2002;9:789–798. ArticlePubMed
- 16. Shyu YJ, Liu H, Deng X, Hu CD. Identification of new fluorescent protein fragments for bimolecular fluorescence complementation analysis under physiological conditions. Biotechniques 2006;40:61–66. ArticlePubMed
- 17. Kallenberger BC, Love JD, Chatterjee VK, Schwabe JW. A dynamic mechanism of nuclear receptor activation and its perturbation in a human disease. Nat Struct Biol 2003;10:136–140. ArticlePubMed
- 18. McKenna NJ, Lanz RB, O'Malley BW. Nuclear receptor coregulators: cellular and molecular biology. Endocr Rev 1999;20:321–344. ArticlePubMed
- 19. Plutzky J. The PPAR-RXR transcriptional complex in the vasculature: energy in the balance. Circ Res 2011;108:1002–1016. ArticlePubMed
- 20. Ziouzenkova O, Plutzky J. Lipolytic PPAR activation: new insights into the intersection of triglycerides and inflammation? Curr Opin Clin Nutr Metab Care 2004;7:369–375. ArticlePubMed
- 21. Han SH, Quon MJ, Koh KK. Beneficial vascular and metabolic effects of peroxisome proliferator-activated receptor-alpha activators. Hypertension 2005;46:1086–1092. ArticlePubMed
References
Fig. 1Identification of a novel variant of mitochondrial trans-2-enoyl-CoA reductase (MECR; cytosolic form of MECR [cMECR]) cDNA. (A) The genomic structure of MECR and cMECR and their coding regions are shown. Inverted triangles indicate the start and stop codons. The dotted-line box indicates the inserted nucleotide sequence. (B) Alignment of the MECR and cMECR sequences. The positions of the upper and lower primers are marked with arrows. (C) Expression patterns of MECR and cMECR in various tissues. The upper polymerase chain reaction band is cMECR, and the lower band is MECR. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as a positive control. Br, brain; He, heart; Ki, kidney; Li, liver; Lu, lung; Pa, pancreas; Sp, spleen; Th, thymus. aEach start codon.
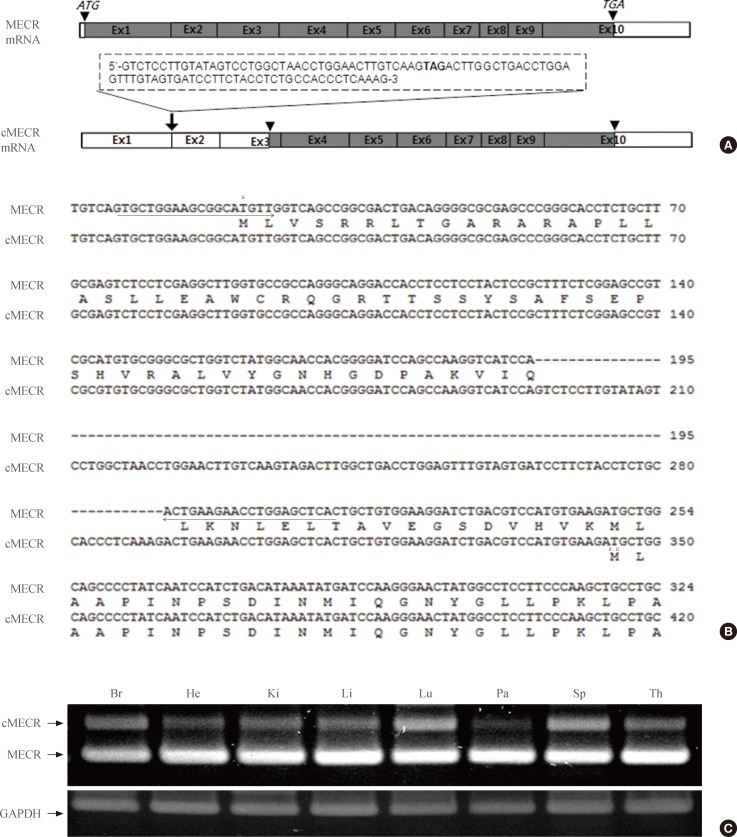
Fig. 2Subcellular localization of mitochondrial trans-2-enoyl-CoA reductase (MECR) and cytosolic form of MERC (cMECR) in HeLa cells. (A) The two constructs used in this study: MECR and cMECR were tagged at the C-terminus with enhanced green fluorescent protein (EGFP; green oval). The black box in MECR indicates the mitochondrial targeting domain. HeLa cells were transiently transfected with (B) MECR-EGFP or (C) cMECR-EGFP and then stained with Mito Tracker, a mitochondrial marker (red). Scale bar=20 µm.
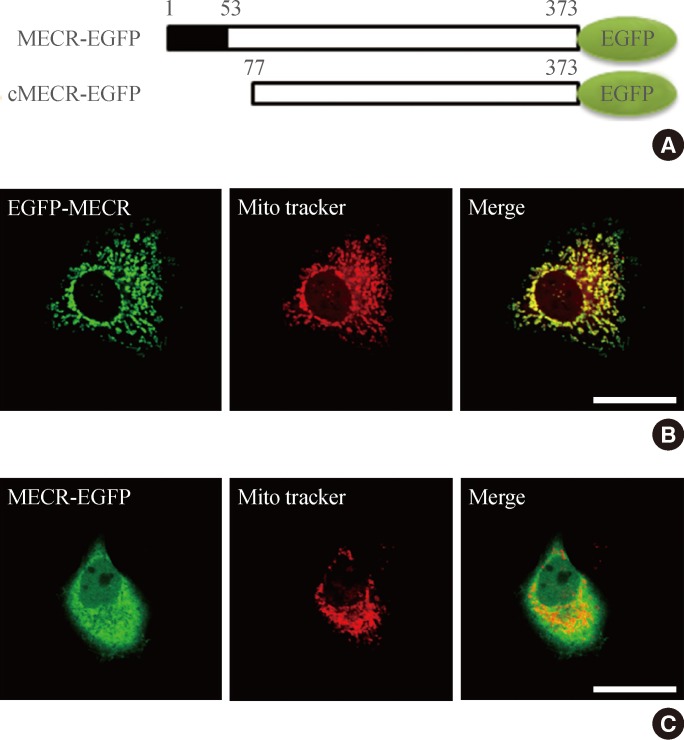
Fig. 3Cytosolic form of mitochondrial trans-2-enoyl-CoA reductase (cMECR) directly interacts with peroxisome proliferator-activated receptor α (PPARα) in vitro. (A) Yeast two-hybrid assay between cMECR and PPARα: on a Trp-/Leu-/His- plate, colonies indicate an interaction between the two genes. (B) Coimmunoprecipitation assays with cMECR and PPARα: cMECR-FLAG was cotransfected with hemagglutinin (HA)-PPARα in HeLa cells. The lysates were immunoprecipitated with anti-FLAG antibody and then immunoblotted with anti-HA antibody. (C) Colocalization of cMECR and PPARα: HeLa cells were transiently transfected with cMECR-enhanced green fluorescent protein (EGFP; green) and mCherry-PPARα (red). Scale bar=20 µm.
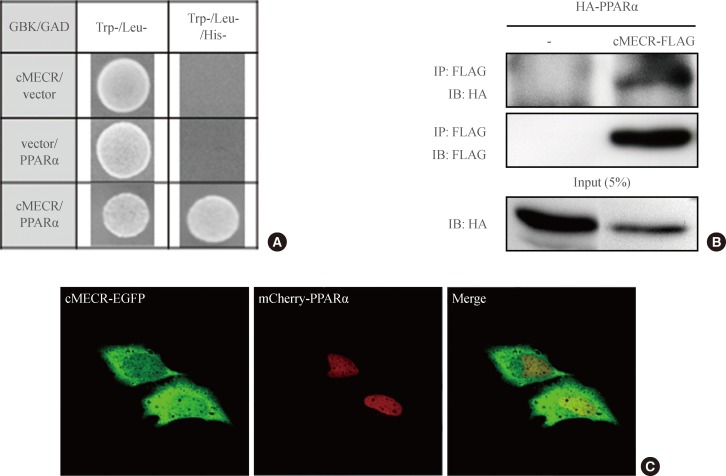
Fig. 4Cytosolic form of mitochondrial trans-2-enoyl-CoA reductase (cMECR) potentiates peroxisome proliferator-activated receptor α (PPARα) activity. (A) Schematic diagram to explain the use of the venus fusion proteins for bimolecular fluorescence complementation (BiFC). (B) BiFC of cMECR with PPARα: plasmids containing cMECR-N terminal fragment of venus (VN) and C terminal fragment of venus (VC)-PPARα were cotransfected into HeLa cells. These cells were stained with 4',6-diamidino-2-phenylindole (DAPI). Venus fluorescence is pseudocolored green. Scale bar=20 mm. (C) Luciferase assay of PPARα: PPARα activity was measured in HeLa cells expressing the PPAR-Luciferase reporter plasmid, HA-PPARα, and cMECR-FLAG. All values are mean±s.e.m. P values were obtained with Student's t test. aP<0.01.

Supplementary Fig. 1(A) Nucleotide sequence showing exon/intron junctions for the generation of cytosolic form of mitochondrial trans-2-enoyl-CoA reductase (cMECR): all exon-intron boundaries conform to consensus splice-donor and splice-acceptor sites (GT-AG). (B) Amino acid alignment of rat cMECR with human MECR isoform 2: the amino acid sequences of both proteins were aligned using the Uniprot alignment program. Sequence identity is 87.2%. Asterisks mean same amino acid.

Supplementary Fig. 2(A) Schematic drawing of the effect of enhanced green fluorescent protein (EGFP) tags on the localization of mitochondrial trans-2-enoyl-CoA reductase (MECR). (B) N-terminal EGFP-tagged MECR is mostly primarily located in cytosolic regions. C-terminal EGFP-tagged MECR localizes to mitochondrial regions. HeLa cells were transiently transfected with EGFP-MECR or MECR-EGFP. Scale bar=20 µm.

Supplementary Fig. 3Localization of mitochondrial trans-2-enoyl-CoA reductase (MECR) and peroxisome proliferator-activated receptor α (PPARα): wild-type MECR is localized in cytosolic regions, whereas PPARα is localized in nuclear regions. HeLa cells were transiently transfected with MECR-enhanced green fluorescent protein (EGFP; green) and mCherry-PPARα (red). Scale bar=20 µm.

Figure & Data
References
Citations
Citations to this article as recorded by 
- Metabolism of phenolics in coffee and plant-based foods by canonical pathways: an assessment of the role of fatty acid β-oxidation to generate biologically-active and -inactive intermediates
Michael N. Clifford, Laurence J. King, Asimina Kerimi, Maria Gema Pereira-Caro, Gary Williamson
Critical Reviews in Food Science and Nutrition.2024; 64(11): 3326. CrossRef - Comparison of muscle nutritional composition, texture quality, carotenoid metabolites and transcriptome to underling muscle quality difference between wild-caught and pond-cultured Yellow River carp (Cyprinus carpio haematopterus)
Luming Wang, Jinrui Xiong, Chunchu Xu, Chaobin Qin, Yuru Zhang, Liping Yang, Shaoyang Zhi, Jianxin Feng, Guoxing Nie
Aquaculture.2024; 581: 740392. CrossRef - Effects of microcystin-LR on immune function, lipid metabolism and intestinal microbial structure in Eriocheir sinensis
Jinliang Du, Liping Cao, Jiancao Gao, Zhijuan Nie, Quanjie Li, Yi Sun, Nailin Shao, Jiawen Hu, Lin Zhou, Guojun Yin, Gangchun Xu
Aquaculture Reports.2024; 35: 101994. CrossRef - A defect in mitochondrial fatty acid synthesis impairs iron metabolism and causes elevated ceramide levels
Debdeep Dutta, Oguz Kanca, Seul Kee Byeon, Paul C. Marcogliese, Zhongyuan Zuo, Rishi V. Shridharan, Jun Hyoung Park, Guang Lin, Ming Ge, Gali Heimer, Jennefer N. Kohler, Matthew T. Wheeler, Benny A. Kaipparettu, Akhilesh Pandey, Hugo J. Bellen
Nature Metabolism.2023; 5(9): 1595. CrossRef - Alternative splicing liberates a cryptic cytoplasmic isoform of mitochondrial MECR that antagonizes influenza virus
Steven F. Baker, Helene Meistermann, Manuel Tzouros, Aaron Baker, Sabrina Golling, Juliane Siebourg Polster, Mitchell P. Ledwith, Anthony Gitter, Angelique Augustin, Hassan Javanbakht, Andrew Mehle, Frank Kirchhoff
PLOS Biology.2022; 20(12): e3001934. CrossRef - Genetic variants in ALDH1L1 and GLDC influence the serine-to-glycine ratio in Hispanic children
Sergey A Krupenko, Shelley A Cole, Ruixue Hou, Karin Haack, Sandra Laston, Nitesh R Mehta, Anthony G Comuzzie, Nancy F Butte, V Saroja Voruganti
The American Journal of Clinical Nutrition.2022; 116(2): 500. CrossRef - Simultaneous Presentation of Multiple Myeloma and Lung Cancer: Case Report and Gene Bioinformatics Analysis
Ping-Ping Xiao, Bing-Qing Luo, Wei Fan, Xu-Yan Chen, Zhi-Gao Dong, Jin-Mei Huang, Yi Zhang, Yong-Quan Chen
Frontiers in Oncology.2022;[Epub] CrossRef - Fatty acid metabolism-related genes are associated with flavor-presenting aldehydes in Chinese local chicken
Xiaoya Yuan, Huanxian Cui, Yuxi Jin, Wenjuan Zhao, Xiaojing Liu, Yongli Wang, Jiqiang Ding, Li Liu, Jie Wen, Guiping Zhao
Frontiers in Genetics.2022;[Epub] CrossRef - NRBF2-mediated autophagy contributes to metabolite replenishment and radioresistance in glioblastoma
Jeongha Kim, Hyunkoo Kang, Beomseok Son, Min-Jung Kim, JiHoon Kang, Kang Hyun Park, Jaewan Jeon, Sunmi Jo, Hae Yu Kim, HyeSook Youn, BuHyun Youn
Experimental & Molecular Medicine.2022; 54(11): 1872. CrossRef - Mitochondrial Fatty Acids and Neurodegenerative Disorders
Alexander J. Kastaniotis, Kaija J. Autio, Remya R. Nair
The Neuroscientist.2021; 27(2): 143. CrossRef - The effects of chronic cadmium exposure on Bufo gargarizans larvae: Histopathological impairment, gene expression alteration and fatty acid metabolism disorder in the liver
Zongqi Ju, Jing Ya, Xinyi Li, Hongyuan Wang, Hongfeng Zhao
Aquatic Toxicology.2020; 222: 105470. CrossRef - Exploration of targets regulated by miR-125b in porcine adipocytes
Xiao Cheng, Xingping Chen, Peng Wang, Ting Chen, Jiajie Sun, Qianyun Xi, Yongliang Zhang
In Vitro Cellular & Developmental Biology - Animal.2020; 56(2): 103. CrossRef - Mitochondrial fatty acid synthesis coordinates oxidative metabolism in mammalian mitochondria
Sara M Nowinski, Ashley Solmonson, Scott F Rusin, J Alan Maschek, Claire L Bensard, Sarah Fogarty, Mi-Young Jeong, Sandra Lettlova, Jordan A Berg, Jeffrey T Morgan, Yeyun Ouyang, Bradley C Naylor, Joao A Paulo, Katsuhiko Funai, James E Cox, Steven P Gygi,
eLife.2020;[Epub] CrossRef - Polymorphisms in ten candidate genes are associated with conformational and locomotive traits in Spanish Purebred horses
Natalia Sevane, Susana Dunner, Ana Boado, Javier Cañon
Journal of Applied Genetics.2017; 58(3): 355. CrossRef - Deep RNA sequencing of pectoralis muscle transcriptomes during late-term embryonic to neonatal development in indigenous Chinese duck breeds
Chunhong Zhu, Weitao Song, Zhiyun Tao, Hongxiang Liu, Wenjuan Xu, Shuangjie Zhang, Huifang Li, Cristina Óvilo
PLOS ONE.2017; 12(8): e0180403. CrossRef - Mitochondrial fatty acid synthesis, fatty acids and mitochondrial physiology
Alexander J. Kastaniotis, Kaija J. Autio, Juha M. Kerätär, Geoffray Monteuuis, Anne M. Mäkelä, Remya R. Nair, Laura P. Pietikäinen, Antonina Shvetsova, Zhijun Chen, J. Kalervo Hiltunen
Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids.2017; 1862(1): 39. CrossRef - Genetic modifications of Mecr reveal a role for mitochondrial 2-enoyl-CoA/ACP reductase in placental development in mice
Remya R. Nair, Juha M. Kerätär, Kaija J. Autio, Ali J. Masud, Mikko A.J. Finnilä, Helena I. Autio-Harmainen, Ilkka J. Miinalainen, Pentti A. Nieminen, J. Kalervo Hiltunen, Alexander J. Kastaniotis
Human Molecular Genetics.2017; 26(11): 2104. CrossRef - MECR Mutations Cause Childhood-Onset Dystonia and Optic Atrophy, a Mitochondrial Fatty Acid Synthesis Disorder
Gali Heimer, Juha M. Kerätär, Lisa G. Riley, Shanti Balasubramaniam, Eran Eyal, Laura P. Pietikäinen, J. Kalervo Hiltunen, Dina Marek-Yagel, Jeffrey Hamada, Allison Gregory, Caleb Rogers, Penelope Hogarth, Martha A. Nance, Nechama Shalva, Alvit Veber, Mic
The American Journal of Human Genetics.2016; 99(6): 1229. CrossRef - Genome‐wide association study with the risk of schizophrenia in a Korean population
Lyoung Hyo Kim, Byung Lae Park, Hyun Sub Cheong, Suhg Namgoong, Ji On Kim, Jeong‐Hyun Kim, Joong‐Gon Shin, Chul Soo Park, Bong‐Jo Kim, Jae Won Kim, Ihn‐Geun Choi, Jaeuk Hwang, Hyoung Doo Shin, Sung‐Il Woo
American Journal of Medical Genetics Part B: Neuropsychiatric Genetics.2016; 171(2): 257. CrossRef - A global transcriptional analysis of Megalobrama amblycephala revealing the molecular determinants of diet-induced hepatic steatosis
Dingdong Zhang, Kangle Lu, Guangzhen Jiang, Wenbin Liu, Zaijie Dong, Hongyan Tian, Xiangfei Li
Gene.2015; 570(2): 255. CrossRef - Articles in 'Endocrinology and Metabolism' in 2014
Won-Young Lee
Endocrinology and Metabolism.2015; 30(1): 47. CrossRef
 PubReader
PubReader Cite
Cite