The Role of Nuclear Factor-E2-Related Factor 1 in the Oxidative Stress Response in MC3T3-E1 Osteoblastic Cells
Article information

Abstract
Background
Reactive oxygen species (ROS) and antioxidants are associated with maintenance of cellular function and metabolism. Nuclear factor-E2-related factor 1 (NFE2L1, Nrf1) is known to regulate the expression of a number of genes involved in oxidative stress and inflammation. The purpose of this study was to examine the effects of NFE2L1 on the response to oxidative stress in osteoblastic MC3T3-E1 cells.
Methods
The murine calvaria-derived MC3T3-E1 cell line was exposed to lipopolysaccharide (LPS) for oxidative stress induction. NFE2L1 effects were evaluated using small interfering RNA (siRNA) for NFE2L1 mRNA. ROS generation and the levels of known antioxidant enzyme genes were assayed.
Results
NFE2L1 expression was significantly increased 2.4-fold compared to the control group at 10 µg/mL LPS in MC3T3-E1 cells (P<0.05). LPS increased formation of intracellular ROS in MC3T3-E1 cells. NFE2L1 knockdown led to an additional increase of ROS (20%) in the group transfected with NFE2L1 siRNA compared with the control group under LPS stimulation (P<0.05). RNA interference of NFE2L1 suppressed the expression of antioxidant genes including metallothionein 2, glutamatecysteine ligase catalytic subunit, and glutathione peroxidase 1 in LPS-treated MC3T3-E1 cells.
Conclusion
Our results suggest that NFE2L1 may have a distinct role in the regulation of antioxidant enzymes under inflammation-induced oxidative stress in MC3T3-E1 osteoblastic cells.
INTRODUCTION
Reactive oxygen species (ROS) are considered to be a causal factor in inflammation, aging and a number of degenerative diseases such as atherosclerosis, carcinogenesis, infarction, and osteoporosis [1]. The effects of ROS are eliminated by enzymatic mechanisms involved in cellular antioxidant defense and xenobiotic detoxification [2]. The delicate balance between ROS and antioxidants is important to maintain equilibrium between osteoblasts and osteoclasts activities, respectively, under physiological conditions [3]. ROS and antioxidants are also known to be involved in the pathogenesis of bone loss such as in osteoporosis [45].
Lipopolysaccharide (LPS) is a constituent of the cell wall outer membrane of gram-negative bacteria and has various biological effects including immune and inflammatory responses [6]. LPS has the capacity to induce bone resorption in vitro and also stimulates osteoblasts to secrete osteolytic factors [7]. LPS is involved in the suppression of bone sialoprotein, a mineralized tissue-specific protein in osteoblast-like ROS 17/2.8 cells [8].
Nuclear factor-E2-related factor 1 (NFE2L1, Nrf1) is a basic leucine zipper protein (bZIP) in the Cap-N-Collar (CNC) transcriptional factor family and controls antioxidant response element (ARE)-driven genes [9]. NFE2L1 was originally suggested to have a role in β-globin gene expression in erythroid cells; however, NFE2L1 has also been shown to bind the ARE and regulate the expression of many genes involved in oxidative stress, cellular differentiation, and inflammation [10].
NFE2L1 can protect cells from oxidative stress by regulating genes encoding enzymes related to glutathione (GSH) biosynthesis and other oxidative defense enzymes [10]. Synthesis of GSH, a major antioxidant in the cell, involves γ-glutamylcysteine ligase (γ-GCL), which consists of a catalytic (GCLC) and a modifier (GCLM) light chain [11]. Evidence suggests that NFE2L1 regulates transcription of GSH synthesis-related genes and other antioxidant genes including NAD(P)H dehydrogenase, quinone 1 (NQO1), ferritin-H, metallothionein (MT)-1 and -2 in fibroblasts, and hepatocytes [12131415]. However, despite these observations, there have been no investigations of the function of NFE2L1 in oxidative stress and the NFE2L1-related antioxidant system in osteoblasts.
In the present study, we attempted to assess the effects of inflammation-induced oxidative stress on NFE2L1 expression pattern and also determined a role of NFE2L1 in the expressions of antioxidant-related enzymes using MC3T3-E1 osteoblastic cells.
METHODS
Cell culture and treatment
The murine calvaria-derived MC3T3-E1 osteoblast-like cell line was used in this study. MC3T3-E1 cells were maintained in α-modified minimum essential medium (α-MEM) containing antibiotics and 10% fetal bovine serum. This basic medium was replenished every 3 days.
MC3T3-E1 cells were seeded in either 96- or 6-well plates. Cells were then treated with different concentrations of LPS (Sigma-Aldrich, St. Louis, MO, USA). Cells were subsequently washed twice with phosphate-buffered saline (PBS), and then cells were harvested for experiments.
Transfection of small interfering RNA
MC3T3-E1 cells were plated in either 96- or 6-well plates. After overnight culture, the cells were transfected using Lipofectamine PLUS reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's protocol. Each transfection assay was performed with control small interfering RNA (siRNA) or NFE2L1 siRNA (siNFE2L1) (Santa Cruz Biotechnology, Santa Cruz, CA, USA).
RNA isolation and quantitative real-time polymerase chain reaction
Cultured cells were superficially washed with PBS, followed by extraction of total RNA using Trizol (Invitrogen) according to the manufacturer's standard instructions. Samples (2 µg) of total RNA were reverse transcribed, followed by oligo (dT) primer and MMLV Reverse Transcriptase (Promega Co., Madison, WI, USA) at a final volume of 25 µL. Aliquots of 2 µL cDNA were used as templates for real-time polymerase chain reaction (PCR). PCR amplification was performed with 2×SYBR Premix Ex Ta (Takara Bio Inc., Shiga, Japan) and 10 pmol forward and reverse primers using Thermal Cycler DICE Real Time System (Takara Bio Inc.). Reactions were performed for 45 cycles of 95℃ for 10 seconds, 60℃ for 15 seconds, and 72℃ for 30 seconds. Primers are listed in Table 1.

Primers Used
Measurement of intracellular ROS
Generation of intracellular ROS was measured according to the method described by Wang and Lou [16]. Briefly, MC3T3-E1 cells were cultured on 96-well plates (1×103 cells/well) and transfected with control siRNA (siCONT) or siNFE2L1. After 24 hours, cells were incubated in α-MEM containing fluorescent dye 50 µM H2DCF-DA (Invitrogen) for 15 minutes in the dark, washed thoroughly by PBS, and further incubated in α-MEM with or without 10 µg/mL LPS. The emitted fluorescence was measured by fluorometer (Wallac 1420D Fluorometer, PerkinElmer Inc., Turku, Finland) with excitation and emission wavelengths at 485 and 535 nm, respectively.
Statistical analysis
Statistical analysis was performed with SPSS version 11.5 (SPSS Inc., Chicago, IL, USA). Results are expressed as the mean±SE and the statistical significance was determined by Student t test or one-way analysis of variance with Tukey post hoc test. Significance was defined by a P<0.05.
RESULTS
NFE2L1 expression after LPS treatment
We examined the effect of LPS treatment on NFE2L1 mRNA expression in MC3T3-E1 cells (Fig. 1). Cells were treated with 0, 2, 5, or 10 µg/mL LPS for 24 hours. Although LPS treatment showed a trend to stimulate the expression of NFE2L1 gene in MC3T3-E1 cells dose-dependently, NFE2L1 expression was significantly increased 2.4-fold compared to the control group at 10 µg/mL LPS (P<0.05) (Fig. 1B).

The effect of lipopolysaccharide (LPS) on nuclear factor-E2-related factor 1 (NFE2L1) mRNA expression in MC3T3-E1 cells. Cells were treated with 0, 2, 5, or 10 µg/mL LPS for 24 hours. Levels of mRNA were analyzed by (A) semi-quantitative polymerase chain reaction (PCR) and (B) quantitative real time-PCR. The expression level of each mRNA was normalized to the β-actin levels. aP<0.05 compared with the control group.
RNA interference of NFE2L1
To investigate the contribution of NFE2L1 to the oxidative stress response in MC3T3-E1 cells, specific siRNA for NFE2L1 was transfected in MC3T3-E1 cells. Transfection with siNFE2L1 knocked down expression of NFE2L1 by 79% at 20 nm siRNA as determined by quantitative real time-PCR (P<0.05) (Fig. 2).
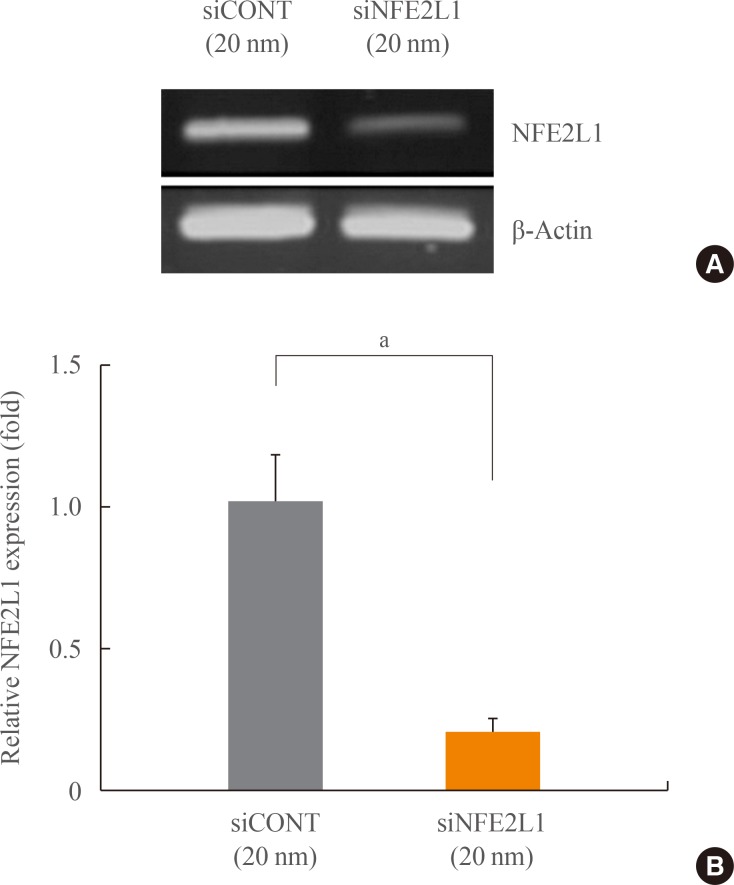
Nuclear factor-E2-related factor 1 (NFE2L1) mRNA expression after transient transfection with control siRNA (siCONT) or NFE2L1 siRNA (siNFE2L1) in MC3T3-E1 cells. Levels of mRNA were analyzed by semi-quantitative (A) polymerase chain reaction (PCR) and (B) quantitative real time-PCR. The expression level of each mRNA was normalized to the β-actin levels. aP<0.05 compared with the siCONT group.
Effects of NFE2L1 knockdown on ROS formation in MC3T3-E1 cells
MC3T3-E1 cells transfected with siCONT or siNFE2L1 were treated with LPS (10 µg/ml) for 10 minutes, and ROS generation was subsequently analyzed (Fig. 3). Quiescent cells (without stimulation) displayed a similar level of ROS in both si-CONT and siNFE2L1 groups. LPS exposure resulted in a significant increase in the amount of ROS by 26% compared with unstimulated cells (P<0.05). The siNFE2L1 led to an additional increase of ROS (20%) compared with the siCONT group under LPS stimulation (P<0.05).
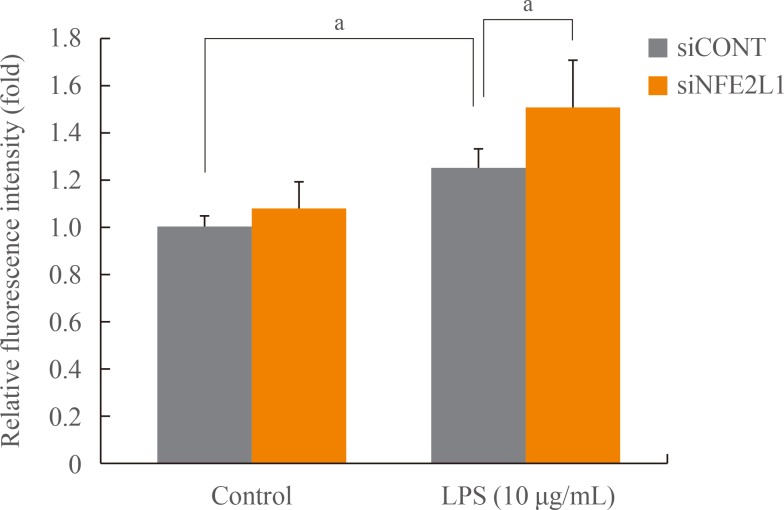
Measurement of reactive oxygen species (ROS) with H2DCF-DA in MC3T3-E1 cells. Intracellular ROS in the transfectants of control siRNA (siCONT) and nuclear factor-E2-related factor 1 siRNA (siNFE2L1) were compared under control (no stimulation) and stimulations by lipopolysaccharide (LPS, 10 µg/mL) for 10 minutes. aP<0.05 compared with the control group or siCONT cells.
Effects of NFE2L1 knockdown on antioxidant gene expression in LPS treated cells
We next examined the effect of mRNA inhibition of NFE2L1 on the mRNA expression of antioxidant genes against oxidative stress in MC3T3-E1 cells. As shown in Fig. 4, there was no significant difference in antioxidant gene expression between siCONT and siNFE2L1 groups under unstimulated conditions. Exposure of MC3T3-E1 cells to LPS led to a significant increase in the level of MT2 compared to untreated controls, and NFE2L1 knockdown resulted in a decrease of 48% in MT2 expression under LPS stimulation (P<0.05) (Fig. 4B). In the presence of LPS, siNFE2L1 also significantly suppressed the expression of GCLC and glutathione peroxidase 1 (GPx1) by 41% and 37%, respectively, compared to the siCONT group (P<0.05) (Fig. 4C, F). However, the differences in mRNA levels of MT1, GCLM, and NQO1 between siCONT and siNFE2L1 groups following stimulation with LPS were not significant.

The effect of nuclear factor-E2-related factor 1 (NFE2L1) knockdown on antioxidant gene mRNA expression in lipopolysaccharide (LPS) treated cells. Antioxidant genes are as follows: (A) metallothionein 1 (MT1), (B) metallothionein 2 (MT2), (C) glutamate-cysteine ligase catalytic subunit (GCLC), (D) glutamate-cysteine ligase modifier subunit (GCLM), (E) NAD(P)H dehydrogenase, quinone 1 (NQO1), and (F) glutathione peroxidase 1 (GPx1). MC3T3-E1 cells were transfected with control siRNA (siCONT) or NFE2L1 siRNA (siNFE2L1) followed by 24-hour treatment of 10 µg/mL LPS. Controls received culture medium only. Quantitation of mRNA levels was analyzed by quantitative real-time polymerase chain reaction. The expression level of each mRNA was normalized to the β-actin levels. aP<0.05 compared with the control group or siCONT cells.
DISCUSSION
Bone remodeling is controlled by a wide range of systemic factors including hormones and steroids and local factors as well as bacterial products such as LPSs. Bacteria-induced pathological bone remodeling is related to bacterial arthritis, osteitis, osteomyelitis, and periodontitis [7]. Bone loss at sites of infection results from mainly increased formation and function of boneresorbing osteoclasts, though bacterially stimulated osteoblasts showed the ability to produce considerable inflammatory mediators that can promote osteoclastogenesis [17].
NFE2L1 is a member of the CNC family of bZIP transcriptional factors and plays an important role in the control of a wide range of genes involved in antioxidants, differentiation, and inflammation [10]. While an understanding of the role of NFE2L1 in the stress response has been demonstrated in various cells, it remains unclear whether NFE2L1 has unique functions under different conditions.
Here we found that expression of NFE2L1 was increased dose-dependently by LPS in MC3T3-E1 cells. LPS is known to have biologic effects to stimulate the production of cytokines such as interleukin 1 (IL-1), IL-6, and tumor necrosis factor α from osteoblasts [7] and these factors contribute to produce ROS in nonphagocytic cells [18]. We then observed that ROS generation by LPS was additionally increased after NFE2L1 silencing. Other studies have reported that NFE2L1-deficient hepatocytes and fibroblasts showed elevated free radicals under normal conditions and with cytotoxic agents, respectively [1219]. Our results suggest that osteoblasts could deal with the increased intracellular oxidative stress burden partially mediated by NFE2L1.
In this study, LPS treatment led to a strong induction of MT2 expression in MC3T3-E1 cells and functional inhibition of NFE2L1 by siRNA caused a significant decrease in expression of MT2 in the presence of LPS. The MT gene is known to be regulated transcriptionally in vivo by heavy metals, glucocorticoid hormones, and LPS [202122]. The level of MT2 expression is relatively higher than MT1, with the ratio of MT1 mRNA to MT2 mRNA ranging from 2:3 to 5:7 [23]. The mouse MT1 and MT2 genes are located in close proximity on chromosome 8 and are amplified together by heavy metals such as cadmium [2324]. Although the ARE of the mouse MT1 gene is preferentially regulated by NFE2L1 [15], the presence of two forms of MT genes could allow greater flexibility in the regulation of expression depending on the different types of inducers. Therefore, our study suggests that LPS might act as a strong inducer of MT2 expression in osteoblastic MC3T3-E1 cells, which is dominantly affected by NFE2L1 under oxidative stress.
We also observed that the expression of GCLC and GPx1 was affected after NFE2L1 knockdown under LPS stimulation. The mRNA expression of GPx1 and GCLC genes might be upregulated to a negligible extent with LPS-only treatment. Li et al. [25] reported that the levels of GPx were not changed in LPS-stimulated macrophages but significantly elevated in LPS-activated cells exposed to an antioxidative stress reagent. Previous studies have shown that GCLC and GPx1 were induced in response to overexpression of NFE2L1 or increased intracellular accumulation of NFE2L1 [1426]. The expression of GCLM and NQO1 was not changed by LPS and knockdown of NFE2L1, as NFE2L2 has been shown to primarily regulate GCLM and NQO1 [2728].
Our study has several limitations. First, oxidative stress could be induced by many agents such as H2O2, high glucose, cytotoxic drugs and metal; however, we focused on inflammation-induced oxidative stress using LPS. Second, we did not assess the association of NFE2L1 with forkhead homeobox type O-1, which is a crucial regulator of oxidative stress in osteoblasts [29]. Third, NFE2L1 is known to involve in cellular differentiation [3031]. It remains to be examined whether osteoblasts differentiation is affected by the change of NFE2L1 expression and antioxidant enzymes under LPS-induced oxidative stress. Finally, we only included the knock-down system of target gene, not performing over-expression system in this study.
In conclusion, this is the first study to elucidate the effects of NFE2L1 on inflammation-induced oxidative stress and the response of antioxidant enzymes in MC3T3-E1 cells. Our results show that the NFE2L1 gene is induced by LPS treatment and NFE2L1 mediates expression of antioxidant enzymes under oxidative stress induced by LPS in osteoblastic MC3T3-E1 cells. This work suggests that NFE2L1 has a distinct function in regulating the response to oxidative stress in osteoblastic cells.
ACKNOWLEDGMENTS
This study was funded by the Korean Endocrine Society.
Notes
CONFLICTS OF INTEREST: No potential conflict of interest relevant to this article was reported.
