Articles
- Page Path
- HOME > Endocrinol Metab > Volume 31(1); 2016 > Article
-
Review ArticleUnderstanding Metabolomics in Biomedical Research
-
Su Jung Kim1, Su Hee Kim1, Ji Hyun Kim1, Shin Hwang2, Hyun Ju Yoo1

-
Endocrinology and Metabolism 2016;31(1):7-16.
DOI: https://doi.org/10.3803/EnM.2016.31.1.7
Published online: March 16, 2016
1Biomedical Research Center, Department of Convergence Medicine, Asan Institute for Life Sciences, Asan Medical Center, University of Ulsan College of Medicine, Seoul, Korea.
2Division of Liver Transplantation and Hepatobiliary Surgery, Department of Surgery, Asan Medical Center, University of Ulsan College of Medicine, Seoul, Korea.
- Corresponding author: Hyun Ju Yoo. Biomedical Research Center, Department of Convergence Medicine, Asan Institute for Life Sciences, Asan Medical Center, University of Ulsan College of Medicine, 88 Olympic-ro 43-gil, Songpa-gu, Seoul 05505, Korea. Tel: +82-2-3010-4029, Fax: +82-2-3010-8566, yoohyunju@amc.seoul.kr
Copyright © 2016 Korean Endocrine Society
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
ABSTRACT
- The term "omics" refers to any type of specific study that provides collective information on a biological system. Representative omics includes genomics, proteomics, and metabolomics, and new omics is constantly being added, such as lipidomics or glycomics. Each omics technique is crucial to the understanding of various biological systems and complements the information provided by the other approaches. The main strengths of metabolomics are that metabolites are closely related to the phenotypes of living organisms and provide information on biochemical activities by reflecting the substrates and products of cellular metabolism. The transcriptome does not always correlate with the proteome, and the translated proteome might not be functionally active. Therefore, their changes do not always result in phenotypic alterations. Unlike the genome or proteome, the metabolome is often called the molecular phenotype of living organisms and is easily translated into biological conditions and disease states. Here, we review the general strategies of mass spectrometry-based metabolomics. Targeted metabolome or lipidome analysis is discussed, as well as nontargeted approaches, with a brief explanation of the advantages and disadvantages of each platform. Biomedical applications that use mass spectrometry-based metabolomics are briefly introduced.
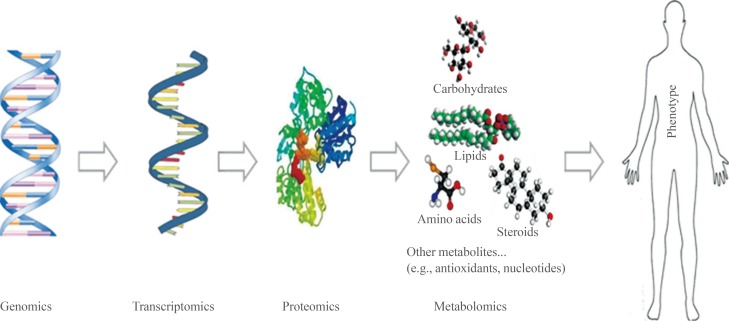
- Metabolites are typically low molecular weight (<1,500 Da) biomolecules. They are the building blocks of the genome, proteome, and cell membranes and play key roles in biology as signaling molecules, energy sources, and metabolic intermediates. Their levels provide integrative information on biological functions and define the phenotypes of biological systems in response to genetic or environmental changes (Fig. 1). Metabolite analysis involves the use of different analytical techniques to identify and quantify intracellular and extracellular metabolites. Successful metabolomic studies rely on proper sample preparation, innovative instrumentation, and bioinformatic tools [1]. Metabolites show broad variations in physicochemical properties, such as polarity, acidity, and volatility. Thus, it is difficult to simultaneously analyze a broad range of metabolites [2].
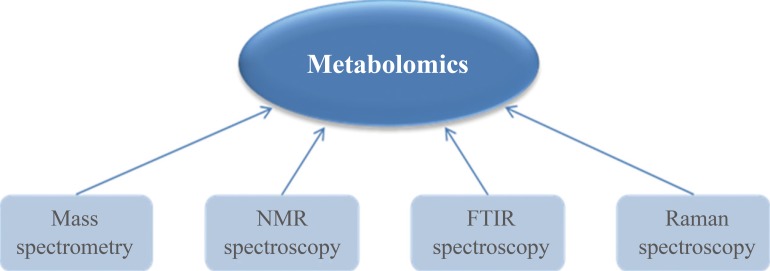
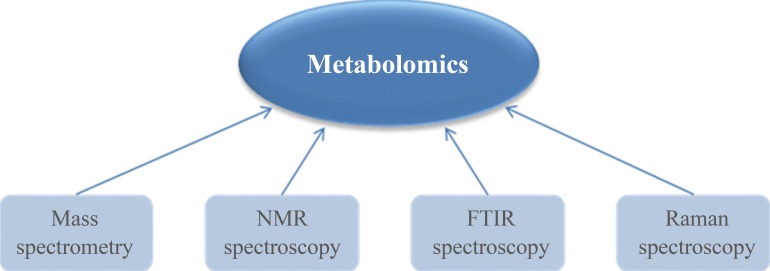
- Metabolomics has developed along with innovative analytical instrumentation. Various analytical techniques have all been applied for metabolite analysis, including mass spectrometry (MS), nuclear magnetic resonance (NMR), Fourier transform infrared (FT-IR) spectroscopy, and Raman spectroscopy (Fig. 2) [3456789]. FT-IR spectroscopy and Raman spectroscopy are rapid, nondestructive, and high-throughput analytical methods that be used for various metabolites. These spectroscopic methods are generally applied to profile metabolic alterations due to their holistic nature, and absorptions at specific wavelengths are sometimes able to provide important clues for the identification of observed metabolic components. However, the sensitivity and selectivity of FT-IR spectroscopy and Raman spectroscopy are lower than those of other methods [8]. NMR spectroscopy, which is a commonly used strategy in metabolomics, is also a rapid and nondestructive method with minimal sample preparation. Chemical shifts in NMR spectra can provide crucial evidence for the identification of specific metabolites. However, NMR requires at least millimolar concentrations of metabolites in samples and its sensitivity is lower than that of MS. Another major disadvantage of NMR is its poor dynamic range [7]. On the other hand, MS remains the most favorable technology for metabolomics due to its wide dynamic range and good sensitivity (nM). In addition, MS is able to detect a diverse range of molecular species comparing to other strategies.
INTRODUCTION
- Recently, MS-based metabolomics became the most popular metabolomics strategy. To establish metabolomic platforms, separation modules are often connected to the MS system. Liquid chromatography/MS (LC/MS) has several advantages over MS alone. Ion suppression, which is often caused by a complex biological matrix, can be eliminated because a metabolite can elute at different retention times with any disturbed metabolites. In addition, the chromatographic peak area provides a useful tool for metabolite quantification because accurate quantification cannot be obtained when MS is used alone.
- Gas chromatography/MS (GC/MS) has been the most suitable MS method for the detection of volatile metabolites. This technique often requires chemical derivatization to improve volatility, and its application has limitation in terms of the molecular size, volatility, and polarity. Therefore, LC/MS has become a popular choice in metabolomics. Sample derivatization is generally not required in LC/MS, and metabolites with more diverse chemical structures and increased molecular sizes can be measured [4810].
- MS-based metabolomics may have difficulties in the identification and characterization of observed metabolites. Mass spectral libraries have not been complete enough for identification of all observed metabolites. The Golm metabolome database consists of mostly plant metabolites, and the National Institute of Standards and Technology database includes only electron ionization-MS data [11]. The METLIN database contains an annotated list of metabolite structural information with MS; however, the database is still not sufficient to identify all observed metabolites [12]. Recently, the Human Metabolome Database (HMDB) was also established to provide chemical and molecular biochemistry data on human metabolites [13]. In addition to the previously mentioned web-based databases, metabolite identification has been reported by commercially available or in-house databases. However, the existence of isobaric or isomeric metabolites may result in misidentification. MS/MS involving collision-induced dissociation has been used to obtain structural information on metabolites [514151617]. The characteristic fragmentation pattern of a metabolite is important for the identification of unknown metabolites and analytical specificity. In addition, the specific retention time in certain separation conditions or the isotopic patterns of unknown metabolites should provide clues for metabolite identification and structural information.
MS-BASED METABOLOMICS
- Nontargeted metabolomics aims to profile the entire metabolome present in cells, biofluids, or tissues. Nontargeted metabolomics measures as many metabolites as possible to compare biological samples and is useful for hypothesis generation. However, the greatest bottleneck in nontargeted metabolomics is the identification of unknown features [18]. Metabolites are found in low picomolar to millimolar concentrations with various physicochemical properties. Thus, it is impossible to observe all metabolites present in a biological sample. Sample preparation and proper analytical instrumentation are important to detect the maximum number of metabolites. High-resolution MS techniques such as time of flight, Orbitrap, or Fourier transform ion cyclotron resonance should be used in nontargeted metabolomics because accurate mass determination is crucial in metabolite identification [1].
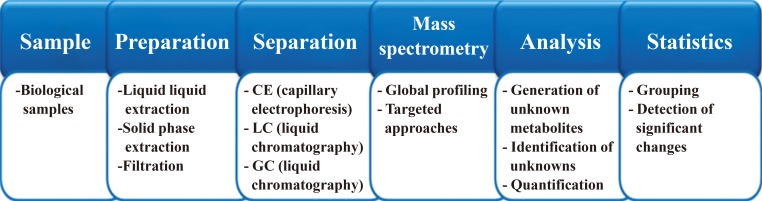
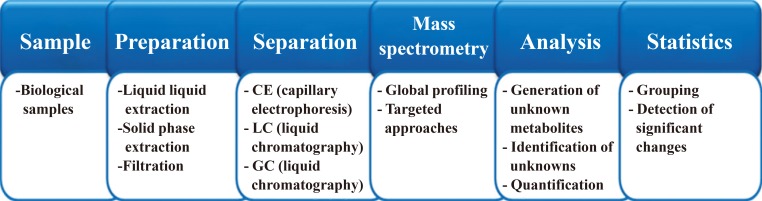
- Global metabolome screening is highly reliant on state-of-the art instruments, bioinformatic tools, and software. General procedures to perform global metabolome profiling involve sample preparation, instrumentation, and statistical or bioinformatic analysis and have been described in several prestigious journals [192021]. General workflow for global metabolome profiling can be found in Fig. 3.
- Sample preparation can be performed to reduce the sample matrix effect and improve sensitivity. Metabolites in a biological sample can sometimes be divided into aliquots of similar physicochemical properties. Liquid-liquid extraction is often used to prepare a sample into two groups of either hydrophobic metabolites or hydrophilic metabolites. Sometimes, simple filtration can be performed to remove solid matter from biological samples, particularly urine. Deproteinization with organic solvents is commonly performed for biofluids.
- Metabolic profiling studies have been performed using various analytical platforms such as capillary electrophoresis (CE)/MS, GC/MS, and LC/MS. Any single analytical platform cannot observe all metabolites in a sample due to the complexity of the concentrations and physicochemical properties of the metabolites. To increase the coverage of observed metabolites, multiple analytical platforms should be used for global metabolome profiling [2223]. GC/MS is usually the preferred platform for small and volatile metabolites, including steroids and fatty acids, and chemical derivatization is often performed to increase the volatility of metabolites. LC/MS can detect a broader range of metabolites, regardless of their hydrophilic or hydrophobic nature, and CE/MS is more appropriate than other platforms for detecting hydrophilic metabolites [24]. Although multiple analytical platforms are recommended for nontargeted metabolomics, not all laboratories have access to all of these analytical instrumentations. Thus, in many cases, the analytical platforms used are chosen depending on their availability and the application.
- Global metabolome profiling data should be acquired from both positive and negative ion mode when the LC/MS or CE/MS platform is used. Positive ion mode can detect basic metabolites, whereas negative ion mode can better detect acidic metabolites. Thus, the maximum coverage of metabolites can be obtained when mass spectrometric data are acquired from both positive and negative ion mode.
- Accordingly, the sample condition and preparation, chromatographic separation, and type of mass analyzer should be considered to obtain high-quality data for global metabolome profiling. The data acquired by global metabolome profiling consists of a list of the mass to charge ratios (m/z) of intact metabolites metabolites and their retention times. Metabolomic software performs data processing and provides such features as P values and fold changes between samples. XCMS and MZmine are freely available web-based interfaces for metabolomic data analysis [252627]. To identify the unknown metabolites of interest, the accurate mass of the metabolite is used to find matches in the metabolite database, such as HMDB and METLIN. Database matches may not be found for many of the observed peaks, and sometimes numerous hits for one peak can be found, even with a narrow range of mass accuracy (often 10 ppm). Correct assignment can be done by comparing the retention time and MS/MS data. Because small molecules can share the same mass and elemental composition, database search results cannot guarantee the correct identification of observed metabolites and the results have to be confirmed with other strategies such as targeted metabolomics. The metabolites identified can be subjected to biological pathway analysis to find relationships with specific disorders or physiological characteristics using web-based databases such as Kyoto Encyclopedia of Genes and Genomes (KEGG), MetaCyc, and BioCyc [282930].
NONTARGETED METABOLOMICS
- Targeted metabolomics provides quantitative information for a predefined list of metabolites or metabolic pathways of interest and is useful for answering any specific biochemical questions or hypotheses [313233]. Each targeted platform is composed of the known metabolites, and the specific analytical method for each platform is established using authentic standards or chemically synthesized compounds. The quantitative information provided by targeted approaches is reliable and has better sensitivity than nontargeted approaches. Without specific analytical platforms, it is often difficult to observe low-level metabolites, namely, those that are present at a low concentration in a biological system. Thus, targeted metabolic platforms are especially preferable for low-level metabolites, which are often bioactive and play important roles in biological systems. Also, when the main interest is a specific class of metabolites or a metabolic pathway, targeted approaches can provide better insight into the specific goal. Targeted metabolic analysis can be a benefit for some metabolites that are not observed well due to their molecular nature, especially when easily ionizable metabolites or isobaric compounds are present.
- It has been reported that the human serum metabolome contains about 20 chemical classes, although other classifications are also possible depending on one's perspective [23]. Different analytical methods have been established to specifically detect each class of metabolite or metabolites related to a metabolic pathway [234]. Commonly, GC or LC separation conditions and different MS/MS techniques can be used to distinguish isobaric or structurally very similar metabolites [35].
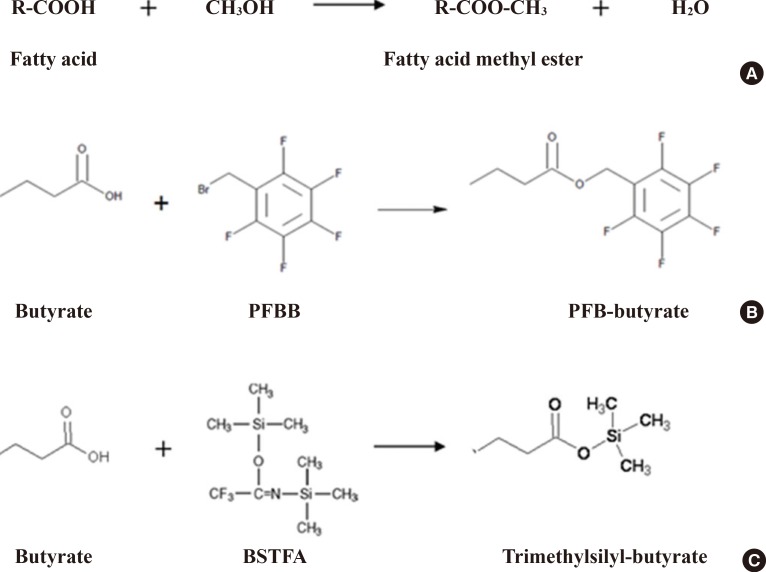
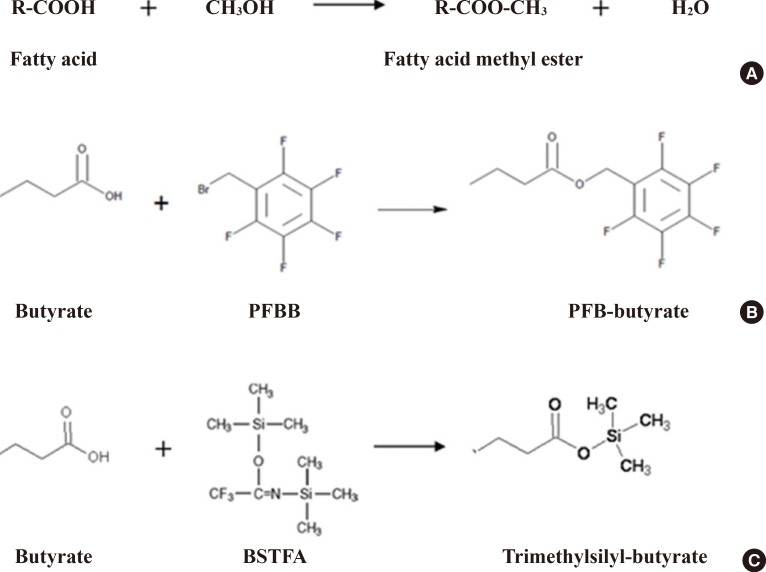
- As an example, targeted analytical platforms of short-chain fatty acids, fatty acids, and eicosanoids will be explained in the following paragraph. Short-chain fatty acids (e.g., acetate, butyrate, propionate, and lactate) are those fatty acids with less than six carbons. They have different chemical and biological properties than fatty acids with more than six carbons. Longer fatty acids are less volatile and less water soluble than short-chain fatty acids. Thus, the extraction protocol should depend on the type of fatty acid. Extraction of short-chain fatty acids from biological samples involves the incubation of the samples with water and uses the aqueous phase for analysis [36]. On the other hand, organic phase extraction should be performed to obtain longer-chain fatty acids from biological samples. Methyl esterification using methanol with BCl3 is a commonly accepted chemical derivatization method for longer-chain fatty acids, but short-chain fatty acids have been derivatized by several methods, such as BSTFA (N, O-bis(trimethyl-silyl) trifluoroacetamide) for trimethylsilylation or PFBB (2,3,4,5,6-pentafluorobenzyl bromide) for PFB (pentafluorobenzyl)-linked derivatives (Fig. 4). These fatty acid derivatives are separated using a GC column, with optimized column type and temperature gradient, then ionized using an electron impact source and analyzed with MS. Intact molecular ions are rarely observed due to the characteristics of the electron impaction source, and specific fragment ions for each fatty acid are used for identification and quantification.
- Eicosanoids are oxidized 20-carbon fatty acids that play key roles in inflammation and immunity. The levels of eicosanoids and their balances affect various biological processes and diseases. Fatty acids in human serum are present at micromolar concentrations, but most eicosanoids in human serum are found at nanomolar or lower concentrations. Thus, the use of solid phase extraction is well established for capturing eicosanoids from complex biological matrixes. Eicosanoids have similar chemical structures, and some of them are isobaric. Accordingly, LC separation is very important to distinguish different eicosanoids among isobaric compounds [37]. Prostaglandin D2 (PGD2) and prostaglandin E2 (PGE2) are isobaric and their elemental compositions are the same. The only difference between PGD2 and PGE2 is their stereochemistry. Careful optimization of LC conditions permits separation of PGD2 and PGE2, as shown in Fig. 5A. About 40 eicosanoids can be analyzed using the LC-MS/MS system, in which the multiple reaction monitoring mode is applied to specifically detect each eicosanoid (Fig. 5B).
- As demonstrated, targeted metabolome platforms should be established to separate target metabolites for better ionization and quantification using sample preparation and a separation module, and to specifically detect metabolites from complex biological matrixes using characteristic fragment ions generated by MS/MS. Greater effort is required in targeted approaches, but targeted metabolome platforms provide absolute or relative quantitative data for specific metabolic pathways once the platform is successfully established.
- Targeted metabolic profiling without standard metabolites is also useful to understand biology or disease states. Phospholipid profiling is a good example. The headgroups of phospholipids are easily observed in positive ion mode during collision activated dissociation, which is a commonly used MS/MS technique, whereas fatty acyl chains are observed in negative ion mode. Thus, LC-MS/MS analysis in both positive and negative ion modes provides structural information on phospholipids. Accordingly, a specific MS/MS technique known as neutral loss scan or precursor ion scan is able to profile a certain class of phospholipids, and relative quantification is also possible by comparing the peak areas of masses corresponding to specific phospholipids.
TARGETED METABOLOMICS
- Metabolites are substrates and products of diverse biochemical reactions and commonly related to the basic biology of the genome and proteome. Metabolomics aims to explore chemical–biological interactions and understand the roles of metabolites in basic, translational, and clinical research. Metabolites have played very important roles in clinical applications, with more than 95% of clinical assays using metabolites [38]. The most representative example would be determination of the blood glucose level for diabetes patients. Most common drugs used today are still small molecules, and more than half of them are derived from metabolites [39]. In addition, human diseases are related to unfavorable interactions of the genome or proteome with metabolites, and a disordered metabolism is involved in many genetic disorders. Recent developments in analytical platforms that use MS have enabled metabolomics to become an important strategy in biomedical research.
- One of the earliest studies using metabolomics in diabetes was plasma phospholipid metabolic profiling for class separation between diabetes mellitus type 2 and the control [40]. This study showed that metabolomics could be used in class separation using multivariate statistical analysis such as partial least square discriminant analysis and principal component analysis and provide potential biomarkers for discrimination. Some groups subsequently performed metabolic studies using both the LC-MS/MS system and H-NMR (proton nuclear magnetic resonance spectroscopy) NMR to explore metabolic pathways altered by insulin deficiency. Two analytical platforms revealed that several metabolic pathways, such as amino acid oxidation, mitochondrial bioenergetics, and gluconeogenesis, were perturbed during insulin deficiency [41]. Newgard et al. [42] used metabolomic profiling to reveal that a branched-chain amino acid-related metabolic signature differentiates obese and lean humans, and contributes to insulin resistance. Through metabolite profiles of human subjects, Wang et al. [43] found five amino acids that can predict an earlier risk of diabetes development. They performed metabolite profiling on the samples of control and prediabetic groups matched according to sex, age, body mass index (BMI), and fasting glucose during a 12-year follow-up period. Branched-chain amino acids (leucine, isoleucine, valine) and aromatic amino acids (phenylalanine, tyrosine) were significantly altered in the prediabetic group. They also measured these five metabolites in a randomly chosen control group that was not matched according to sex, age, BMI, and fasting glucose, confirming the significance of these metabolites. In addition, they assessed the predictive performance of the combination of the five amino acids for the risk of diabetes. In their study, they used a targeted metabolomics strategy, measuring amino acids, urea cycle metabolites, and nucleotide metabolites. On the other hand, Zhao et al. [44] used nontargeted metabolomics to investigate the metabolic signatures of prediabetic subjects in both urine and plasma. They found that fatty acid, tryptophan, uric acid, bile acid, and lysophosphatidylcholine metabolism were the prediabetic-associated alterations.
- Fibrosis develops differentially in different individuals, and there has been no report of any reliable factors that can predict fibrosis susceptibility. Mitochondrial dysfunction has been suggested to be closely related to fibrosis through metabolic perturbations, with Maeda [45] reporting that a radiation-induced fibrosis model had mitochondrial homeostasis problems. Lipid accumulation, including that of ceramides, diacylglycerols, and sphingomyelins, was caused by reduced fatty acid oxidation. This observation led to the possible use of metabolomics to find predictive or diagnostic markers. The plasma fatty acid composition was different in both cystic fibrosis patients and animal models. Bile acids for lipid absorption and cholesterol metabolism were altered in cystic fibrosis, which indicates the imbalanced lipid metabolism in cystic fibrosis [4647]. In addition, sphingosine-1-phosphate was increased due to upregulation of sphingosine kinase in human fibrotic liver [48]. The abnormal lipid metabolism in liver mostly results in inflammation and liver fibrosis [49]. Mice fed a high-fat diet experiences cardiac fibrosis even before obesity and hyperlipidemia develops [50]. Inflammatory lipid mediators such as prostaglandins, leukotrienes, and epoxyeicosatrienoic acids might be involved in cardiac fibrosis [51]. Changes in bioactive phospholipids such as phosphatidic acid and lysophosphatidic acid, compounds involved in cell proliferation, migration, and survival, were observed in the bronchoalveolar lavage of patients suffering from idiopathic pulmonary fibrosis [52]. Branched amino acids have been reported to reduce liver fibrosis via transforming growth factor β inhibition [53]. These studies indicated that a metabolomics study would be a promising strategy for identifying prognostic or diagnostic biomarkers of fibrosis and for better understanding of disease mechanisms.
- Most cancer cells predominantly produce energy for rapid cell proliferation via highly activated glycolysis that is characterized by lactic acid accumulation. This phenomenon of cancer cells, also known as the Warburg effect, indicates an adaptation to the low oxygen environments within tumors or damage of mitochondria in cancer [54]. Metabolic reprogramming in cancer has attracted attention as a new avenue for cancer research [555657]. Metabolic alterations in cancer might include numerous metabolic pathways affecting cell growth and proliferation, including those of energy metabolism, lipid metabolism, and nucleotide metabolism [58]. Bioactive lipids such as eicosanoids are linked to cancer. The roles of these bioactive lipids in the interactions between transformed epithelial cells and the surrounding stromal cells are crucial for understanding tumor evolution, progression, and metastasis [59]. However, the roles of lipids in tumors have been poorly elucidated. The global lipid profiles of the breast cancer tissues of 267 human subjects showed that de novo fatty acid synthesis was higher in tumors than in normal breast tissues, possibly for the incorporation of fatty acids into membrane phospholipids. Immunohistochemical analysis of the expression of specific proteins, selected from in silico transcriptome database searches, showed that lipid metabolism related to de novo fatty acid synthesis was highly expressed [60]. Further understanding of the molecular mechanisms underlying the roles of metabolites in cancer progression should be useful for the development of more effective targets for cancer therapies.
METABOLOMICS APPLICATIONS IN BIOMEDICAL RESEARCH
- Metabolomics is a relatively new omics strategy in biomedical research and is gaining attention as a tool for the discovery of biomarkers for disease diagnosis and assessment of disease development and prognosis. In addition, metabolomics may provide evidence for the identification of candidates for novel therapeutic interventions and the elucidation of disease mechanisms. The successful application of metabolomics requires integrated efforts from analytical chemistry, biology, medicine, and biostatistics.
CONCLUSIONS
-
Acknowledgements
- This study was supported by grants from the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT & Future Planning (2014R1A1A1003608, 2015R1A2A2A04007141).
ACKNOWLEDGMENTS
-
CONFLICTS OF INTEREST: No potential conflict of interest relevant to this article was reported.
Article information
- 1. Fernie AR, Trethewey RN, Krotzky AJ, Willmitzer L. Metabolite profiling: from diagnostics to systems biology. Nat Rev Mol Cell Biol 2004;5:763–769. ArticlePubMedPDF
- 2. Yoo HJ, Liu H, Hakansson K. Infrared multiphoton dissociation and electron-induced dissociation as alternative MS/MS strategies for metabolite identification. Anal Chem 2007;79:7858–7866. ArticlePubMed
- 3. Villas-Boas SG, Mas S, Akesson M, Smedsgaard J, Nielsen J. Mass spectrometry in metabolome analysis. Mass Spectrom Rev 2005;24:613–646. ArticlePubMed
- 4. Want EJ, Cravatt BF, Siuzdak G. The expanding role of mass spectrometry in metabolite profiling and characterization. Chembiochem 2005;6:1941–1951. ArticlePubMed
- 5. Brown SC, Kruppa G, Dasseux JL. Metabolomics applications of FT-ICR mass spectrometry. Mass Spectrom Rev 2005;24:223–231. ArticlePubMed
- 6. Boskey AL, Mendelsohn R. Infrared spectroscopic characterization of mineralized tissues. Vib Spectrosc 2005;38:107–114. ArticlePubMedPMC
- 7. Defernez M, Colquhoun IJ. Factors affecting the robustness of metabolite fingerprinting using 1H NMR spectra. Phytochemistry 2003;62:1009–1017. ArticlePubMed
- 8. Dunn WB, Ellis DI. Metabolomics: current analytical platforms and methodologies. Trends Analyt Chem 2005;24:285–294.Article
- 9. Deleris G, Petibois C. Applications of FT-IR spectrometry to plasma contents analysis and monitoring. Vib Spectrosc 2003;32:129–136.Article
- 10. Metz TO, Zhang Q, Page JS, Shen Y, Callister SJ, Jacobs JM, et al. The future of liquid chromatography-mass spectrometry (LC-MS) in metabolic profiling and metabolomic studies for biomarker discovery. Biomark Med 2007;1:159–185. ArticlePubMedPMC
- 11. Bino RJ, Hall RD, Fiehn O, Kopka J, Saito K, Draper J, et al. Potential of metabolomics as a functional genomics tool. Trends Plant Sci 2004;9:418–425. ArticlePubMed
- 12. Smith CA, O'Maille G, Want EJ, Qin C, Trauger SA, Brandon TR, et al. METLIN: a metabolite mass spectral database. Ther Drug Monit 2005;27:747–751. ArticlePubMed
- 13. Wishart DS, Jewison T, Guo AC, Wilson M, Knox C, Liu Y, et al. HMDB 3.0: the Human Metabolome Database in 2013. Nucleic Acids Res 2013;41(Database issue):D801–D807. ArticlePubMedPDF
- 14. Edwards JL, Chisolm CN, Shackman JG, Kennedy RT. Negative mode sheathless capillary electrophoresis electrospray ionization-mass spectrometry for metabolite analysis of prokaryotes. J Chromatogr A 2006;1106:80–88. ArticlePubMed
- 15. Buchholz A, Takors R, Wandrey C. Quantification of intracellular metabolites in Escherichia coli K12 using liquid chromatographic-electrospray ionization tandem mass spectrometric techniques. Anal Biochem 2001;295:129–137. ArticlePubMed
- 16. Byrd GD, Ogden MW. Liquid chromatographic/tandem mass spectrometric method for the determination of the tobacco-specific nitrosamine metabolite NNAL in smokers' urine. J Mass Spectrom 2003;38:98–107. ArticlePubMed
- 17. Triolo A, Altamura M, Dimoulas T, Guidi A, Lecci A, Tramontana M. In vivo metabolite detection and identification in drug discovery via LC-MS/MS with data-dependent scanning and postacquisition data mining. J Mass Spectrom 2005;40:1572–1582. ArticlePubMed
- 18. Fiehn O, Garvey WT, Newman JW, Lok KH, Hoppel CL, Adams SH. Plasma metabolomics profiles reflective of glucose homeostasis in non-diabetic and type 2 diabetic obese African-American women. PLoS One 2010;5:e15234ArticlePubMedPMC
- 19. Want EJ, Masson P, Michopoulos F, Wilson ID, Theodoridis G, Plumb RS, et al. Global metabolic profiling of animal and human tissues via UPLC-MS. Nat Protoc 2013;8:17–32. ArticlePubMedPDF
- 20. Dunn WB, Broadhurst D, Begley P, Zelena E, Francis-McIntyre S, Anderson N, et al. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat Protoc 2011;6:1060–1083. ArticlePubMedPDF
- 21. Osborn MP, Park Y, Parks MB, Burgess LG, Uppal K, Lee K, et al. Metabolome-wide association study of neovascular age-related macular degeneration. PLoS One 2013;8:e72737ArticlePubMedPMC
- 22. Bouatra S, Aziat F, Mandal R, Guo AC, Wilson MR, Knox C, et al. The human urine metabolome. PLoS One 2013;8:e73076ArticlePubMedPMC
- 23. Psychogios N, Hau DD, Peng J, Guo AC, Mandal R, Bouatra S, et al. The human serum metabolome. PLoS One 2011;6:e16957ArticlePubMedPMC
- 24. Ramautar R, Somsen GW, de Jong GJ. CE-MS in metabolomics. Electrophoresis 2009;30:276–291. ArticlePubMed
- 25. Benton HP, Wong DM, Trauger SA, Siuzdak G. XCMS2: processing tandem mass spectrometry data for metabolite identification and structural characterization. Anal Chem 2008;80:6382–6389. ArticlePubMedPMC
- 26. Tautenhahn R, Patti GJ, Rinehart D, Siuzdak G. XCMS Online: a web-based platform to process untargeted metabolomic data. Anal Chem 2012;84:5035–5039. ArticlePubMedPMC
- 27. Katajamaa M, Miettinen J, Oresic M. MZmine: toolbox for processing and visualization of mass spectrometry based molecular profile data. Bioinformatics 2006;22:634–636. ArticlePubMedPDF
- 28. Krieger CJ, Zhang P, Mueller LA, Wang A, Paley S, Arnaud M, et al. MetaCyc: a multiorganism database of metabolic pathways and enzymes. Nucleic Acids Res 2004;32(Database issue):D438–D442. ArticlePubMedPMCPDF
- 29. Kanehisa M, Goto S. KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res 2000;28:27–30. ArticlePubMedPMCPDF
- 30. Caspi R, Altman T, Billington R, Dreher K, Foerster H, Fulcher CA, et al. The MetaCyc database of metabolic pathways and enzymes and the BioCyc collection of Pathway/Genome Databases. Nucleic Acids Res 2014;42(Database issue):D459–D471. ArticlePubMedPDF
- 31. Yuan M, Breitkopf SB, Yang X, Asara JM. A positive/negative ion-switching, targeted mass spectrometry-based metabolomics platform for bodily fluids, cells, and fresh and fixed tissue. Nat Protoc 2012;7:872–881. ArticlePubMedPMCPDF
- 32. Quehenberger O, Armando AM, Brown AH, Milne SB, Myers DS, Merrill AH, et al. Lipidomics reveals a remarkable diversity of lipids in human plasma. J Lipid Res 2010;51:3299–3305. ArticlePubMedPMC
- 33. Quehenberger O, Dennis EA. The human plasma lipidome. N Engl J Med 2011;365:1812–1823. ArticlePubMedPMC
- 34. Kim SJ, Kim N, Koh EH, Yoo HJ. Identification of ethanolamine plasmalogens from complex lipid mixtures by MS/MS and Ag adduction. Anal Sci 2012;28:1207–1212. ArticlePubMed
- 35. Kim SJ, Back SH, Koh JM, Yoo HJ. Quantitative determination of major platelet activating factors from human plasma. Anal Bioanal Chem 2014;406:3111–3118. ArticlePubMedPDF
- 36. Huda-Faujan N, Abdulamir AS, Fatimah AB, Anas OM, Shuhaimi M, Yazid AM, et al. The impact of the level of the intestinal short chain fatty acids in inflammatory bowel disease patients versus healthy subjects. Open Biochem J 2010;4:53–58. ArticlePubMedPMC
- 37. Yang J, Schmelzer K, Georgi K, Hammock BD. Quantitative profiling method for oxylipin metabolome by liquid chromatography electrospray ionization tandem mass spectrometry. Anal Chem 2009;81:8085–8093. ArticlePubMedPMC
- 38. Tietz NW. Clinical guide to laboratory tests; Philadelphia: WB Saunders; 1995.
- 39. Wishart DS. Metabolomics: applications to food science and nutrition research. Trends Food Sci Technol 2008;19:482–493.Article
- 40. Wang C, Kong H, Guan Y, Yang J, Gu J, Yang S, et al. Plasma phospholipid metabolic profiling and biomarkers of type 2 diabetes mellitus based on high-performance liquid chromatography/electrospray mass spectrometry and multivariate statistical analysis. Anal Chem 2005;77:4108–4116. ArticlePubMed
- 41. Lanza IR, Zhang S, Ward LE, Karakelides H, Raftery D, Nair KS. Quantitative metabolomics by H-NMR and LC-MS/MS confirms altered metabolic pathways in diabetes. PLoS One 2010;5:e10538ArticlePubMedPMC
- 42. Newgard CB, An J, Bain JR, Muehlbauer MJ, Stevens RD, Lien LF, et al. A branched-chain amino acid-related metabolic signature that differentiates obese and lean humans and contributes to insulin resistance. Cell Metab 2009;9:311–326. ArticlePubMedPMC
- 43. Wang TJ, Larson MG, Vasan RS, Cheng S, Rhee EP, McCabe E, et al. Metabolite profiles and the risk of developing diabetes. Nat Med 2011;17:448–453. ArticlePubMedPMCPDF
- 44. Zhao X, Fritsche J, Wang J, Chen J, Rittig K, Schmitt-Kopplin P, et al. Metabonomic fingerprints of fasting plasma and spot urine reveal human pre-diabetic metabolic traits. Metabolomics 2010;6:362–374. ArticlePubMedPMC
- 45. Maeda S. Pathology of experimental radiation pancarditis. II. Correlation between ultrastructural changes of the myocardial mitochondria and succinic dehydrogenase activity in rabbit heart receiving a single dose of X-ray irradiation. Acta Pathol Jpn 1982;32:199–218. PubMed
- 46. Guilbault C, Wojewodka G, Saeed Z, Hajduch M, Matouk E, De Sanctis JB, et al. Cystic fibrosis fatty acid imbalance is linked to ceramide deficiency and corrected by fenretinide. Am J Respir Cell Mol Biol 2009;41:100–106. ArticlePubMed
- 47. Teichgraber V, Ulrich M, Endlich N, Riethmuller J, Wilker B, De Oliveira-Munding CC, et al. Ceramide accumulation mediates inflammation, cell death and infection susceptibility in cystic fibrosis. Nat Med 2008;14:382–391. ArticlePubMedPDF
- 48. Li C, Zheng S, You H, Liu X, Lin M, Yang L, et al. Sphingosine 1-phosphate (S1P)/S1P receptors are involved in human liver fibrosis by action on hepatic myofibroblasts motility. J Hepatol 2011;54:1205–1213. ArticlePubMed
- 49. Moustafa T, Fickert P, Magnes C, Guelly C, Thueringer A, Frank S, et al. Alterations in lipid metabolism mediate inflammation, fibrosis, and proliferation in a mouse model of chronic cholestatic liver injury. Gastroenterology 2012;142:140–151.e12. ArticlePubMed
- 50. Aubin MC, Lajoie C, Clement R, Gosselin H, Calderone A, Perrault LP. Female rats fed a high-fat diet were associated with vascular dysfunction and cardiac fibrosis in the absence of overt obesity and hyperlipidemia: therapeutic potential of resveratrol. J Pharmacol Exp Ther 2008;325:961–968. ArticlePubMed
- 51. Levick SP, Loch DC, Taylor SM, Janicki JS. Arachidonic acid metabolism as a potential mediator of cardiac fibrosis associated with inflammation. J Immunol 2007;178:641–646. ArticlePubMed
- 52. Castelino FV. Lipids and eicosanoids in fibrosis: emerging targets for therapy. Curr Opin Rheumatol 2012;24:649–655. ArticlePubMed
- 53. Cha JH, Bae SH, Kim HL, Park NR, Choi ES, Jung ES, et al. Branched-chain amino acids ameliorate fibrosis and suppress tumor growth in a rat model of hepatocellular carcinoma with liver cirrhosis. PLoS One 2013;8:e77899ArticlePubMedPMC
- 54. Warburg O. On the origin of cancer cells. Science 1956;123:309–314. ArticlePubMed
- 55. Kim YS, Maruvada P. Frontiers in metabolomics for cancer research: Proceedings of a National Cancer Institute workshop. Metabolomics 2008;4:105–113.ArticlePDF
- 56. Kim K, Aronov P, Zakharkin SO, Anderson D, Perroud B, Thompson IM, et al. Urine metabolomics analysis for kidney cancer detection and biomarker discovery. Mol Cell Proteomics 2009;8:558–570. ArticlePubMedPMC
- 57. Griffin JL, Shockcor JP. Metabolic profiles of cancer cells. Nat Rev Cancer 2004;4:551–561. ArticlePubMedPDF
- 58. Kim S, Rhee JK, Yoo HJ, Lee HJ, Lee EJ, Lee JW, et al. Bioinformatic and metabolomic analysis reveals miR-155 regulates thiamine level in breast cancer. Cancer Lett 2015;357:488–497. ArticlePubMed
- 59. Wang D, Dubois RN. Eicosanoids and cancer. Nat Rev Cancer 2010;10:181–193. ArticlePubMedPMCPDF
- 60. Hilvo M, Denkert C, Lehtinen L, Muller B, Brockmoller S, Seppanen-Laakso T, et al. Novel theranostic opportunities offered by characterization of altered membrane lipid metabolism in breast cancer progression. Cancer Res 2011;71:3236–3245. ArticlePubMed
References
Conventional omics studies in biology. There are various types of omics, from genomics to metabolomics, and new omics studies are being constantly added, such as lipidomics or glycomics.
Instrumentation for metabolomics. Analytical instruments for metabolomics should be able to detect various kinds of metabolites present in biological systems. NMR, nuclear magnetic resonance; FTIR, Fourier transform infrared.
General workflow in metabolomics. The metabolomics workflow generally follows the strategy above. However, detailed experimental procedures can be different, especially for targeted metabolomics.
Chemical derivatization for fatty acids. (A) Methyl esterification reaction of fatty acids. (B) pentafluorobenzyl (PFB) derivatization reaction for short-chain fatty acids. (C) Trimethylsilyl derivatization reaction for short-chain fatty acids. PFBB, 2,3,4,5,6-pentafluorobenzyl bromide. BSTFA, N, O-bis(trimethyl-silyl) trifluoroacetamide.
(A) Extracted ion chromatogram for prostaglandin E2 (PGE2) and prostaglandin D2 (PGD2). Liquid chromatography (LC) separation was able to successfully differentiate these isobaric lipids. (B) Total ion chromatogram observed from eicosanoid profiling using LC-mass spectrometry (MS)/MS. cps, counts per second.
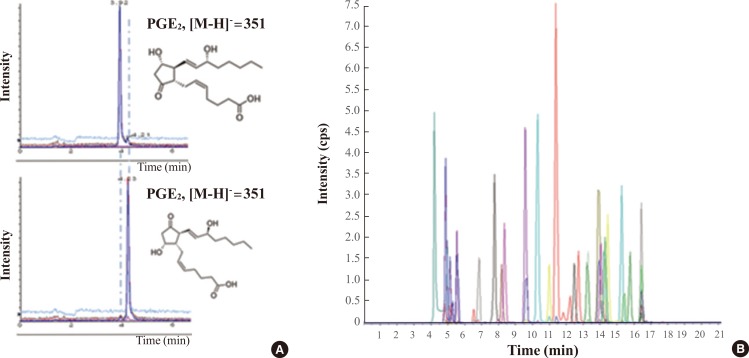
Figure & Data
References
Citations
- Urine metabolomic biomarkers for prediction of isolated fetal congenital heart defect
Perry Friedman, Ali Yilmaz, Zafer Ugur, Faryal Jafar, Amy Whitten, Ilyas Ustun, Onur Turkoglu, Stewart Graham, Ray Bahado Singh
The Journal of Maternal-Fetal & Neonatal Medicine.2024; 35(25): 6380. CrossRef - Metabolomics and Risk of Dementia: A Systematic Review of Prospective Studies
Ashley C Flores, Xinyuan Zhang, Penny M Kris-Etherton, Martin J Sliwinski, Greg C Shearer, Xiang Gao, Muzi Na
The Journal of Nutrition.2024; 154(3): 826. CrossRef - Associations of PFAS-related plasma metabolites with cholesterol and triglyceride concentrations
T. Schillemans, I.A. Bergdahl, K. Hanhineva, L. Shi, C. Donat-Vargas, J. Koponen, H. Kiviranta, R. Landberg, A. Åkesson, C. Brunius
Environmental Research.2023; 216: 114570. CrossRef - UPLC-Q-Exactive-based rats serum metabolomics for characterization of traditional Chinese medicine Natures and Flavors
Hong Wang, Ruofang Gao, Jing Liu, Shuang Zhang, Yunli Zhao, Zhiguo Yu
Journal of Ethnopharmacology.2023; 302: 115931. CrossRef - Artificial Neural Networks Coupled with MALDI-TOF MS Serum Fingerprinting To Classify and Diagnose Pathological Pain Subtypes in Preclinical Models
Meritxell Deulofeu, Eladia M. Peña-Méndez, Petr Vaňhara, Josef Havel, Lukáš Moráň, Lukáš Pečinka, Anna Bagó-Mas, Enrique Verdú, Victoria Salvadó, Pere Boadas-Vaello
ACS Chemical Neuroscience.2023; 14(2): 300. CrossRef - Metabolic signature of HepaRG cells exposed to ethanol and tumor necrosis factor alpha to study alcoholic steatohepatitis by LC–MS-based untargeted metabolomics
Elias Iturrospe, Rani Robeyns, Katyeny Manuela da Silva, Maria van de Lavoir, Joost Boeckmans, Tamara Vanhaecke, Alexander L. N. van Nuijs, Adrian Covaci
Archives of Toxicology.2023; 97(5): 1335. CrossRef - Integrating metabolomics and network pharmacology to assess the effects of quercetin on lung inflammatory injury induced by human respiratory syncytial virus
Ya-Lei Sun, Pei-Pei Zhao, Cheng-Bi Zhu, Ming-Chen Jiang, Xin-Min Li, Jia-Lei Tao, Chan-Chan Hu, Bin Yuan
Scientific Reports.2023;[Epub] CrossRef - Comparison of Plasma Metabolites From Patients With Non-Small Cell Lung Cancer by Erlotinib Treatment and Skin Rash
Won Kil Lee, Jisoo Myong, Eunbin Kwag, Younmin Shin, Ji Woong Son, Byong Chul Yoo, Byoung-Soo Kim, Hwa-Seung Yoo, Jeong June Choi
Integrative Cancer Therapies.2023;[Epub] CrossRef - Metabolomic Signatures of Exposure to Nitrate and Trihalomethanes in Drinking Water and Colorectal Cancer Risk in a Spanish Multicentric Study (MCC-Spain)
Jose A. Alcolea, Carolina Donat-Vargas, Anastasia Chrysovalantou Chatziioannou, Pekka Keski-Rahkonen, Nivonirina Robinot, Antonio José Molina, Pilar Amiano, Inés Gómez-Acebo, Gemma Castaño-Vinyals, Lea Maitre, Marc Chadeau-Hyam, Sonia Dagnino, Sibo Lucas
Environmental Science & Technology.2023; 57(48): 19316. CrossRef - Metabolic Signature of Ethanol-Induced Hepatotoxicity in HepaRG Cells by Liquid Chromatography–Mass Spectrometry-Based Untargeted Metabolomics
Elias Iturrospe, Katyeny Manuela da Silva, Rani Robeyns, Maria van de Lavoir, Joost Boeckmans, Tamara Vanhaecke, Alexander L.N. van Nuijs, Adrian Covaci
Journal of Proteome Research.2022; 21(4): 1153. CrossRef - Current State and Challenges of the Global Outcomes of Dental Caries Research in the Meta-Omics Era
Dina G. Moussa, Paras Ahmad, Tamer A. Mansour, Walter L. Siqueira
Frontiers in Cellular and Infection Microbiology.2022;[Epub] CrossRef - Current Knowledge in Skin Metabolomics: Updates from Literature Review
Alessia Paganelli, Valeria Righi, Elisabetta Tarentini, Cristina Magnoni
International Journal of Molecular Sciences.2022; 23(15): 8776. CrossRef - Metabolome-wide association study of the relationship between chlorpyrifos exposure and first trimester serum metabolite levels in pregnant Thai farmworkers
Donghai Liang, Jonathan Batross, Nancy Fiedler, Tippawan Prapamontol, Panrapee Suttiwan, Parinya Panuwet, Warangkana Naksen, Brittney O. Baumert, Volha Yakimavets, Youran Tan, Priya D'Souza, Ampica Mangklabruks, Supattra Sittiwang, Kristsanachai Kaewthit,
Environmental Research.2022; 215: 114319. CrossRef - Differential Plasma Metabolites between High- and Low-Grade Meningioma Cases
Gabriel A. Kurokawa, Pedro T. Hamamoto Filho, Jeany Delafiori, Aline F. Galvani, Arthur N. de Oliveira, Flávia L. Dias-Audibert, Rodrigo R. Catharino, Maria Inês M. C. Pardini, Marco A. Zanini, Estela de O. Lima, Adriana C. Ferrasi
International Journal of Molecular Sciences.2022; 24(1): 394. CrossRef - Plasma metabolites associated with exposure to perfluoroalkyl substances and risk of type 2 diabetes – A nested case-control study
Tessa Schillemans, Lin Shi, Carolina Donat-Vargas, Kati Hanhineva, Andreas Tornevi, Ingegerd Johansson, Jani Koponen, Hannu Kiviranta, Olov Rolandsson, Ingvar A. Bergdahl, Rikard Landberg, Agneta Åkesson, Carl Brunius
Environment International.2021; 146: 106180. CrossRef - Methionine transsulfuration pathway is upregulated in long-lived humans
Natàlia Mota-Martorell, Mariona Jové, Consuelo Borrás, Rebeca Berdún, Èlia Obis, Joaquim Sol, Rosanna Cabré, Irene Pradas, José Daniel Galo-Licona, Josep Puig, José Viña, Reinald Pamplona
Free Radical Biology and Medicine.2021; 162: 38. CrossRef - Investigating metabolism by mass spectrometry: From steady state to dynamic view
Matteo Audano, Silvia Pedretti, Simona Ligorio, Flavio Giavarini, Donatella Caruso, Nico Mitro
Journal of Mass Spectrometry.2021;[Epub] CrossRef - The Efficacy of Pilates on Urinary Incontinence in Korean Women: A Metabolomics Approach
Gyumin Kang, Haelim Lee, Malsoon Shin, Jaekwan Kim, Sungki Lee, Youngja Park
Metabolites.2021; 11(2): 118. CrossRef - Broad Metabolome Alterations Associated with the Intake of Oral Contraceptives Are Mediated by Cortisol in Premenopausal Women
Clara Eick, Johanna Klinger-König, Stephanie Zylla, Anke Hannemann, Kathrin Budde, Ann Kristin Henning, Maik Pietzner, Matthias Nauck, Henry Völzke, Hans J. Grabe, Johannes Hertel
Metabolites.2021; 11(4): 193. CrossRef - Nutritional Metabolomics and the Classification of Dietary Biomarker Candidates: A Critical Review
Talha Rafiq, Sandi M Azab, Koon K Teo, Lehana Thabane, Sonia S Anand, Katherine M Morrison, Russell J de Souza, Philip Britz-McKibbin
Advances in Nutrition.2021; 12(6): 2333. CrossRef - MSCAT: A Machine Learning Assisted Catalog of Metabolomics Software Tools
Jonathan Dekermanjian, Wladimir Labeikovsky, Debashis Ghosh, Katerina Kechris
Metabolites.2021; 11(10): 678. CrossRef - Defining NASH from a Multi-Omics Systems Biology Perspective
Lili Niu, Karolina Sulek, Catherine G. Vasilopoulou, Alberto Santos, Nicolai J. Wewer Albrechtsen, Simon Rasmussen, Florian Meier, Matthias Mann
Journal of Clinical Medicine.2021; 10(20): 4673. CrossRef - Defining Blood Plasma and Serum Metabolome by GC-MS
Olga Kiseleva, Ilya Kurbatov, Ekaterina Ilgisonis, Ekaterina Poverennaya
Metabolites.2021; 12(1): 15. CrossRef - Precision Medicine in Childhood Asthma: Omic Studies of Treatment Response
Javier Perez-Garcia, Esther Herrera-Luis, Fabian Lorenzo-Diaz, Mario González, Olaia Sardón, Jesús Villar, Maria Pino-Yanes
International Journal of Molecular Sciences.2020; 21(8): 2908. CrossRef - Metabolomic characterization of semen from asthenozoospermic patients using ultra‐high‐performance liquid chromatography–tandem quadrupole time‐of‐flight mass spectrometry
Li Li, Xinrui Hao, Hua Chen, Liyuan Wang, Aolei Chen, Xiaotian Song, Zhiyan Hu, Ying Su, Han Lin, Pei Fan
Biomedical Chromatography.2020;[Epub] CrossRef - Comparison of Metabolomic Profiles of Organs in Mice of Different Strains Based on SPME-LC-HRMS
Katarzyna Burlikowska, Iga Stryjak, Joanna Bogusiewicz, Bogumiła Kupcewicz, Karol Jaroch, Barbara Bojko
Metabolites.2020; 10(6): 255. CrossRef - Metabolomics and Machine Learning Approaches Combined in Pursuit for More Accurate Paracoccidioidomycosis Diagnoses
Estela de Oliveira Lima, Luiz Claudio Navarro, Karen Noda Morishita, Camila Mika Kamikawa, Rafael Gustavo Martins Rodrigues, Mohamed Ziad Dabaja, Diogo Noin de Oliveira, Jeany Delafiori, Flávia Luísa Dias-Audibert, Marta da Silva Ribeiro, Adriana Pardini
mSystems.2020;[Epub] CrossRef - Metabolomics in the study of spontaneous animal diseases
Helena Tran, Malcolm McConville, Panayiotis Loukopoulos
Journal of Veterinary Diagnostic Investigation.2020; 32(5): 635. CrossRef - Effect of Pioglitazone on Perihematomal Edema in Intracerebral Hemorrhage Mouse Model by Regulating NLRP3 Expression and Energy Metabolism
Hoon Kim, Jung Eun Lee, Hyun Ju Yoo, Jae Hoon Sung, Seung Ho Yang
Journal of Korean Neurosurgical Society.2020; 63(6): 689. CrossRef - Inborn Errors of Metabolism in the Era of Untargeted Metabolomics and Lipidomics
Ismail, Showalter, Fiehn
Metabolites.2019; 9(10): 242. CrossRef - Metabolomic profiles of induced pluripotent stem cells derived from patients with rheumatoid arthritis and osteoarthritis
Juryun Kim, Sunyoung Christina Kang, Na Eun Yoon, Yena Kim, Jinhyeok Choi, Narae Park, Hyerin Jung, Byung Hwa Jung, Ji Hyeon Ju
Stem Cell Research & Therapy.2019;[Epub] CrossRef - Metabolomics profiles of patients with Wilson disease reveal a distinct metabolic signature
Gaurav V. Sarode, Kyoungmi Kim, Dorothy A. Kieffer, Noreene M. Shibata, Tomas Litwin, Anna Czlonkowska, Valentina Medici
Metabolomics.2019;[Epub] CrossRef - Metabolomics study of the therapeutic mechanism of a Chinese herbal formula on collagen-induced arthritis mice
Zhen Jin, Ji-da Zhang, Xin Wu, Gang Cao
RSC Advances.2019; 9(7): 3716. CrossRef - Plasma sphingomyelins increase in pre-diabetic Korean men with abdominal obesity
Seung-Soon Im, Hyeon Young Park, Jong Cheol Shon, In-Sung Chung, Ho Chan Cho, Kwang-Hyeon Liu, Dae-Kyu Song, Kyoung Heon Kim
PLOS ONE.2019; 14(3): e0213285. CrossRef - Tissue‐Specific Metabolic Profiles After Prolonged Cardiac Arrest Reveal Brain Metabolome Dysfunction Predominantly After Resuscitation
Jaewoo Choi, Muhammad Shoaib, Tai Yin, Gautam Nayyar, Koichiro Shinozaki, Jan F. Stevens, Lance B. Becker, Junhwan Kim
Journal of the American Heart Association.2019;[Epub] CrossRef - Global serum metabolomics profiling of colorectal cancer (Review)
Nurul Amir Hashim, Sharaniza Ab‑Rahim, Leny Suddin, Mohd Ahmad Saman, Musalmah Mazlan
Molecular and Clinical Oncology.2019;[Epub] CrossRef - Metabolomic Profiling of Human Spermatozoa in Idiopathic Asthenozoospermia Patients Using Gas Chromatography-Mass Spectrometry
Kai Zhao, Jianzhong Zhang, Zhen Xu, Yue Xu, Aiming Xu, Wei Chen, Chenkui Miao, Shouyong Liu, Zengjun Wang, Ruipeng Jia
BioMed Research International.2018; 2018: 1. CrossRef - Articles inEndocrinology and Metabolismin 2016
Won-Young Lee
Endocrinology and Metabolism.2017; 32(1): 62. CrossRef - Investigation of the effects of storage and freezing on mixes of heavy‐labeled metabolite and amino acid standards
Rachel Culp‐Hill, Julie A. Reisz, Kirk C. Hansen, Angelo D'Alessandro
Rapid Communications in Mass Spectrometry.2017; 31(23): 2030. CrossRef - Beyond the paradigm: Combining mass spectrometry and nuclear magnetic resonance for metabolomics
Darrell D. Marshall, Robert Powers
Progress in Nuclear Magnetic Resonance Spectroscopy.2017; 100: 1. CrossRef - Untargeted metabolomics analysis of adipogenic transformation in OP9‐DL1 cells using liquid chromatography‐mass spectrometry: Implications for thymic adipogenesis
Jianxin Tan, Yajun Wang, Siliang Wang, Nannan Zhang, Simeng Wu, Zhe Yuan, Xike Zhu
Cell Biology International.2017; 41(4): 447. CrossRef - Gut Microbiota Profiling: Metabolomics Based Approach to Unravel Compounds Affecting Human Health
Pamela Vernocchi, Federica Del Chierico, Lorenza Putignani
Frontiers in Microbiology.2016;[Epub] CrossRef
 PubReader
PubReader Cite
Cite