Articles
- Page Path
- HOME > Endocrinol Metab > Volume 32(1); 2017 > Article
-
Review ArticleRecent Insights into Insulin-Like Growth Factor Binding Protein 2 Transcriptional Regulation
-
Minsang Shin1
 , Hye Suk Kang2, Jae-Hyung Park2, Jae-Hoon Bae2, Dae-Kyu Song2, Seung-Soon Im2
, Hye Suk Kang2, Jae-Hyung Park2, Jae-Hoon Bae2, Dae-Kyu Song2, Seung-Soon Im2 -
Endocrinology and Metabolism 2017;32(1):11-17.
DOI: https://doi.org/10.3803/EnM.2017.32.1.11
Published online: January 19, 2017
1Department of Microbiology, Kyungpook National University School of Medicine, Daegu, Korea.
2Department of Physiology, Keimyung University School of Medicine, Daegu, Korea.
- Corresponding author: Seung-Soon Im. Department of Physiology, Keimyung University School of Medicine, 1095 Dalgubeol-daero, Dalseo-gu, Daegu 42601, Korea. Tel: +82-53-580-3863, Fax: +82-53-580-3795, ssim73@kmu.ac.kr
• Received: November 18, 2016 • Revised: December 13, 2016 • Accepted: December 20, 2016
Copyright © 2017 Korean Endocrine Society
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
ABSTRACT
- Insulin-like growth factor binding proteins (IGFBPs) are major regulators of insulin-like growth factor bioavailability and activity in metabolic signaling. Seven IGFBP family isoforms have been identified. Recent studies have shown that IGFBPs play a pivotal role in metabolic signaling and disease, including the pathogenesis of obesity, diabetes, and cancer. Although many studies have documented the various roles played by IGFBPs, transcriptional regulation of IGFBPs is not well understood. In this review, we focus on the regulatory mechanisms of IGFBP gene expression, and we summarize the findings of transcription factor activity in the IGFBP promoter region.
- Insulin-like growth factor binding proteins (IGFBPs) are a superfamily member of homologous proteins present in serum [1]. All members of the IGFBP family include a cysteine-rich domain, which contains the GCGCCXXC motif in the N-terminal and C-terminal domains [2]. Insulin-like growth factors (IGFs) are central hormones involved in metabolic signaling, affecting glucose uptake, lipogenesis, glycogen storage, and suppression of protein degradation [3]. Studies have shown that IGFBPs have IGF-dependent and IGF-independent functions [1]. Within the IGFBP family, IGFBP-2 is a protein encoded by the IGFBP-2 gene [4]. Observations on the identification and function of IGFBP-2 in metabolic signaling and disease are discussed in this review. These data provide new insights into our understanding of the pathophysiology of metabolic syndrome and have important clinical implications, although additional research is required.
INTRODUCTION
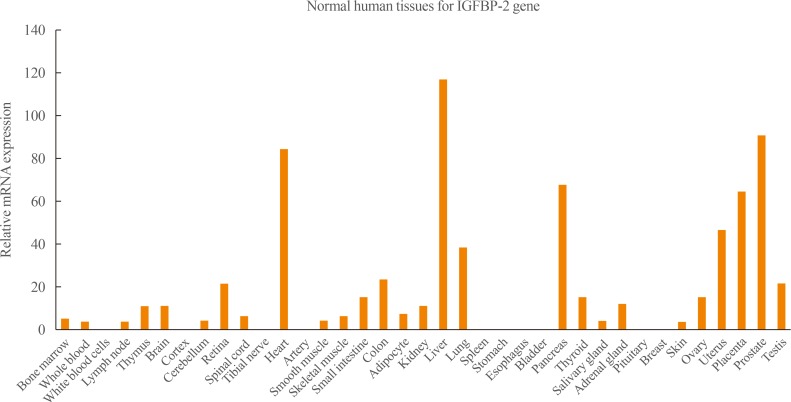
- IGFBP-2 is a 36-kDa protein that it is mainly expressed during embryonic development; it is also expressed in adult liver and adipocytes, and in the central nervous and reproductive systems [5]. Tissue survey data have elucidated the IGFBP-2 gene expression profile (Fig. 1). These data show that IGFBP-2 is mainly expressed in the liver, heart, kidney, and prostate.
TISSUE SURVEY OF IGFBP-2 GENE EXPRESSION
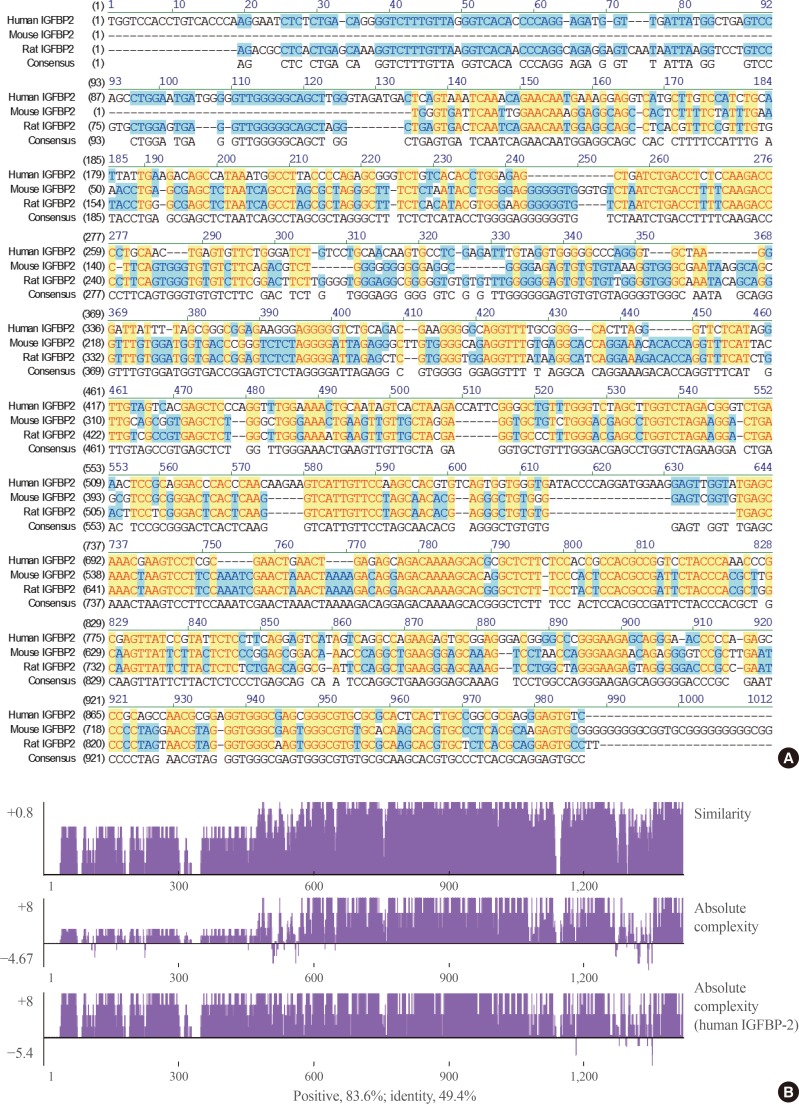
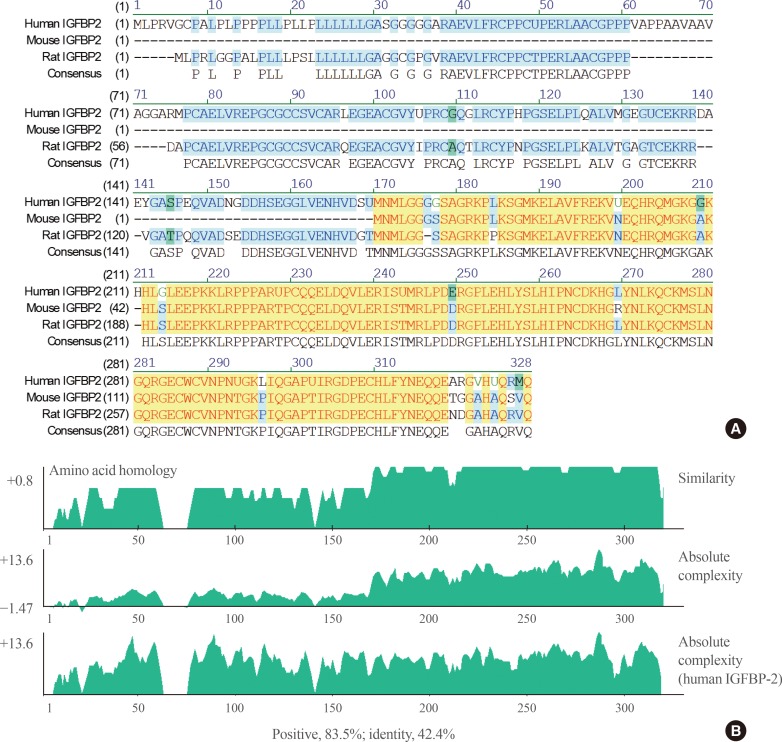
- When the IGFBP-2 sequence was compared between species, several regions of high similarity between promoter sequences were observed (Fig. 2). The IGFBP-2 gene is identical in 49.4% of sequences between humans, rats, and mice. Similarly, exon sequences for the IGFBP-2 gene are about 42.2% identical across species (Fig. 3). In particular, consensus sequences for the major transcription factors match significantly between humans, mice, and rats. Moreover, the IGFBP-2 gene C-terminal domain, including the consensus sequence for IGF binding and the CWCV (cysteine-tryptophan-cysteine-valine) domain, is highly conserved across a variety of species [6]. These data indicate that IGFBP-2 may have high binding affinity with IGFs.
COMPARISON OF IGFBP-2 GENE SEQUENCES BETWEEN SPECIES
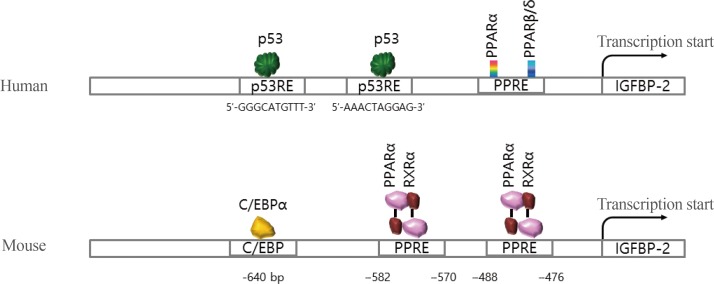
- Although the secretion of IGFBP-2 protein is well established in metabolic signaling, the molecular mechanisms of the IGFBP-2 gene have only recently been elucidated. A few transcription factors, including peroxisome proliferation-activator receptor α (PPARα), multiple endocrine neoplasia type 1 (MEN1), CCAAT-enhancer-binding protein α (C/EBPα), and hypoxia-inducible factor 1 (HIF-1), have been identified on the IGFBP-2 promoter (Fig. 4).
- One study found that IGFBP-2 expression increased during prolonged fasting, and the plasma concentration of IGFBP-2 was consistently induced for 48 hours with no change postprandial or after glucose challenge [7]. Increased IGFBP-2 expression during fasting is regulated by PPARα [89]. The PPARα response element (PPRE) was identified on a mouse IGFBP-2 promoter, indicating that IGFBP-2 is a direct target of PPARα. Moreover, mRNA expression of the IGFBP-2 gene is activated by metformin, which is an antidiabetic drug for patients with diabetes and obesity, and the serum level of IGFBP-2 is stimulated in metformin-challenged diabetic patients [10]. A recent study in humans suggested that the circulating concentration of IGFBP-2 is associated with metabolic syndrome, including insulin resistance and obesity [1112]. These results suggest that overexpression of the IGFBP-2 gene plays a protective role against insulin resistance.
- Previous studies found that insulin suppresses induction of IGFBP-2, as well as IGFBP-1 [13]. Moreover, hepatic IGFBP-2 expression is decreased by insulin infusion, whereas IGFBP-1 mRNA recovers to baseline within 1 hour postinsulin challenge. These results suggest that IGFBP-2 expression level is very important for modulating insulin and IGF-1 sensitivity. In addition, the expression level of IGFBP-2 could potentially serve as a biomarker of metabolic disease, such as diabetes or insulin resistance. Although IGFBP-2 is mainly expressed in the liver, transcriptional regulation of the IGFBP-2 gene has been studied in other tissues [111415]. In particular, IGFBP-2 gene expression in 3T3-L1 adipocytes is regulated by C/EBPα, even though IGFBP-2 is expressed at a very low level in white adipose tissue [16]. Compared to the liver, insulin stimulated IGFBP-2 gene expression in adipocytes. Insulin activates IGFBP-2 expression in 3T3-L1 adipocytes and MCF-7 (Michigan Cancer Foundation 7) breast cancer cell lines through the phosphoinositide 3-kinase/mechanistic target of rapamycin signaling axis [1617]. These findings indicate that insulin signaling might increase IGFBP-2 secretion by activating insulin-induced, adipocyte-specific expression of the C/EBPα gene in differentiated 3T3-L1 adipocytes.
- Transcriptional regulation of the IGFBP-2 gene also occurs in tumorigenesis [8181920]. During tumor growth, IGFBP-2 expression is regulated by the transcription factor HIF-1α in hypoxic conditions [8]. In addition, IGFBP-2 mRNA is upregulated by the tumor suppressor gene p53 in a dose-dependent manner, indicating that IGFBP-2 is a p53 target [20]. Also, the IGFBP-2 gene was upregulated by disruption of the menin gene in an animal model and stable cell lines [2122]. Menin is a nuclear protein-encoded Men1 gene, which is mutated in patients with MEN1 [22]. MEN1 is a syndrome characterized by tumorigenesis in multiple endocrine organs (e.g., pancreas and parathyroids) [22]. Menin is required to repress endogenous IGFBP-2 expression by modifying the chromatin structure surrounding the IGFBP-2 gene promoter. Although IGFBP-2 promoter activity is decreased by overexpression of menin, the regulatory mechanism by which menin affects the IGFBP-2 promoter remains unknown.
TRANSCRIPTIONAL REGULATION OF IGFBP-2
- In a transgenic mouse model, overexpression of IGFBP-2 was associated with increased fat accumulation and reduced muscle weight [23]. Although IGFBP-2 is a secondary protein among the circulation binding proteins, the regulatory and functional roles of IGFBP-2 are not well understood compared to those of IGFBP-1. A recent study determined the role of IGFBP-2 to be a pleiotropic oncogenic protein in cancer development [24]. IGFBP-2 stimulates the nuclear form of epidermal growth factor receptor (EGFR), increasing the activated transcription factor 3 (STAT3) pathway. Moreover, overexpression of exogenous IGFBP-2 protein inhibits EGFR activity through suppression of nuclear EGFR signaling [24]. These results demonstrate a strong association between IGFBP-2 and STAT3 and suggest a novel tumor-inducing role for IGFBP2 by providing a linkage between IGFBP-2 and cancer development.
- A novel function of IGFBP-2 has been reported in Duchenne muscular dystrophy (DMD) [14]. One study injected IGFBP-2 into the muscles of dystrophic mice and found that muscle-specific upregulation of IGFBP-2 inhibited muscular dystrophy and protected against dystrophic pathophysiology. Although these discoveries could eventually lead to gene therapy for DMD, it is not yet possible to directly address the muscle atrophy resulting from neuromuscular disorders such as DMD. IGF-1 signaling slows progression of DMD [252627], and studies have found that overexpression of IGFBP-2 can assist in dystrophic treatment by reducing muscle composition in a muscle-dystrophic animal model and patients with DMD.
FUNCTION OF IGFBPS IN METABOLIC SIGNALING
- Chronic hyperinsulinemia reduces IGFBP-2 secretion, resulting in increased availability of IGF-1 [28]. Studies have shown that IGFBPs serve a critical function in the metabolic system and represent an important link between the IGF system and insulin signaling [29]. In recent years, in vivo and in vitro assays, as well as studies in human subjects, have focused on the functional progression of IGFBPs. These investigations have confirmed the role of IGFBPs in metabolic syndrome, including fatty liver, insulin resistance, and obesity [30]. Production of IGFBP-2 is decreased in obese people compared with nonobese individuals, and concentrations of serum IGFBP-2 decrease with increasing body mass index [13]. Several reports have demonstrated that circulating IGFBP-2 concentrations are lower in patients with type 2 diabetes mellitus or obesity [1031]. Because overexpression of IGFBP-2 leads to recovered blood glucose levels in diabetic animal models, and because the antidiabetic drug, metformin, can recover serum IGFBP-2 levels in diabetic patients [10], decreased IGFBP-2 levels may lead to the development of diabetes in humans. Thus, IGFBP-2 may be useful as a biomarker of metabolic disease.
CLINICAL RELEVANCE OF IGFBP-2 LEVELS IN HUMAN SERUM
- IGFBP-2 has been shown to be a key regulator of metabolic diseases, such as diabetes and obesity, as well as cancer metabolism. Although IGFBP-2 gene transcription is regulated by several metabolic conditions in endocrine organs, the underlying regulatory mechanism of IGFBP-2 gene expression remains unknown, and further studies are needed. Moreover, the functional roles of IGFBP-2 in metabolic diseases and signaling are still unclear and controversial. Therefore, the relevance of IGFBP-2 to metabolic diseases could be a main driver in the investigation of IGFBP-2's underlying physiological function.
CONCLUSIONS
-
Acknowledgements
- This study was supported by grants from the Korea New Faculty, Korea Research Foundation, Republic of Korea (NRF-2013R1A1A1006606) to S.S.I. and Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2015R1D1A1A01057610) to H.S.K.
ACKNOWLEDGMENTS
-
CONFLICTS OF INTEREST: No potential conflict of interest relevant to this article was reported.
Article information
- 1. Wheatcroft SB, Kearney MT. IGF-dependent and IGF-independent actions of IGF-binding protein-1 and -2: implications for metabolic homeostasis. Trends Endocrinol Metab 2009;20:153–162. ArticlePubMed
- 2. Kim HS, Nagalla SR, Oh Y, Wilson E, Roberts CT Jr, Rosenfeld RG. Identification of a family of low-affinity insulin-like growth factor binding proteins (IGFBPs): characterization of connective tissue growth factor as a member of the IGFBP superfamily. Proc Natl Acad Sci U S A 1997;94:12981–12986. ArticlePubMedPMC
- 3. Clemmons DR. The relative roles of growth hormone and IGF-1 in controlling insulin sensitivity. J Clin Invest 2004;113:25–27. ArticlePubMedPMC
- 4. Schoen TJ, Mazuruk K, Waldbillig RJ, Potts J, Beebe DC, Chader GJ, et al. Cloning and characterization of a chick embryo cDNA and gene for IGF-binding protein-2. J Mol Endocrinol 1995;15:49–59. ArticlePubMed
- 5. Shimasaki S, Ling N. Identification and molecular characterization of insulin-like growth factor binding proteins (IGFBP-1, -2, -3, -4, -5 and -6). Prog Growth Factor Res 1991;3:243–266. ArticlePubMed
- 6. Kibbey MM, Jameson MJ, Eaton EM, Rosenzweig SA. Insulin-like growth factor binding protein-2: contributions of the C-terminal domain to insulin-like growth factor-1 binding. Mol Pharmacol 2006;69:833–845. ArticlePubMed
- 7. Clemmons DR, Snyder DK, Busby WH Jr. Variables controlling the secretion of insulin-like growth factor binding protein-2 in normal human subjects. J Clin Endocrinol Metab 1991;73:727–733. ArticlePubMedPDF
- 8. Feldser D, Agani F, Iyer NV, Pak B, Ferreira G, Semenza GL. Reciprocal positive regulation of hypoxia-inducible factor 1alpha and insulin-like growth factor 2. Cancer Res 1999;59:3915–3918. PubMed
- 9. Kang HS, Kim MY, Kim SJ, Lee JH, Kim YD, Seo YK, et al. Regulation of IGFBP-2 expression during fasting. Biochem J 2015;467:453–460. ArticlePubMedPMCPDF
- 10. Kang HS, Cho HC, Lee JH, Oh GT, Koo SH, Park BH, et al. Metformin stimulates IGFBP-2 gene expression through PPARalpha in diabetic states. Sci Rep 2016;6:23665ArticlePubMedPMCPDF
- 11. Li Z, Picard F. Modulation of IGFBP2 mRNA expression in white adipose tissue upon aging and obesity. Horm Metab Res 2010;42:787–791. ArticlePubMedPDF
- 12. Ruan W, Lai M. Insulin-like growth factor binding protein: a possible marker for the metabolic syndrome? Acta Diabetol 2010;47:5–14. ArticlePubMedPDF
- 13. Frystyk J. Free insulin-like growth factors: measurements and relationships to growth hormone secretion and glucose homeostasis. Growth Horm IGF Res 2004;14:337–375. ArticlePubMed
- 14. Swiderski K, Martins KJ, Chee A, Trieu J, Naim T, Gehrig SM, et al. Skeletal muscle-specific overexpression of IGFBP-2 promotes a slower muscle phenotype in healthy but not dystrophic mdx mice and does not affect the dystrophic pathology. Growth Horm IGF Res 2016;30-31:1–10. ArticlePubMed
- 15. Johnson MH, de Mejia EG. Phenolic compounds from fermented berry beverages modulated gene and protein expression to increase insulin secretion from pancreatic beta-cells in vitro. J Agric Food Chem 2016;64:2569–2581. ArticlePubMed
- 16. Li Z, Miard S, Laplante M, Sonenberg N, Picard F. Insulin stimulates IGFBP-2 expression in 3T3-L1 adipocytes through the PI3K/mTOR pathway. Mol Cell Endocrinol 2012;358:63–68. ArticlePubMed
- 17. Mireuta M, Darnel A, Pollak M. IGFBP-2 expression in MCF-7 cells is regulated by the PI3K/AKT/mTOR pathway through Sp1-induced increase in transcription. Growth Factors 2010;28:243–255. ArticlePubMed
- 18. Dunlap SM, Celestino J, Wang H, Jiang R, Holland EC, Fuller GN, et al. Insulin-like growth factor binding protein 2 promotes glioma development and progression. Proc Natl Acad Sci U S A 2007;104:11736–11741. ArticlePubMedPMC
- 19. Tombolan L, Orso F, Guzzardo V, Casara S, Zin A, Bonora M, et al. High IGFBP2 expression correlates with tumor severity in pediatric rhabdomyosarcoma. Am J Pathol 2011;179:2611–2624. ArticlePubMedPMC
- 20. Grimberg A, Coleman CM, Shi Z, Burns TF, MacLachlan TK, Wang W, et al. Insulin-like growth factor factor binding protein-2 is a novel mediator of p53 inhibition of insulin-like growth factor signaling. Cancer Biol Ther 2006;5:1408–1414. ArticlePubMedPMC
- 21. Pannett AA, Thakker RV. Multiple endocrine neoplasia type 1. Endocr Relat Cancer 1999;6:449–473. ArticlePubMed
- 22. La P, Schnepp RW, Petersen CD, Silva AC, Hua X. Tumor suppressor menin regulates expression of insulin-like growth factor binding protein 2. Endocrinology 2004;145:3443–3450. ArticlePubMedPMCPDF
- 23. Rehfeldt C, Renne U, Sawitzky M, Binder G, Hoeflich A. Increased fat mass, decreased myofiber size, and a shift to glycolytic muscle metabolism in adolescent male transgenic mice overexpressing IGFBP-2. Am J Physiol Endocrinol Metab 2010;299:E287–E298. ArticlePubMed
- 24. Chua CY, Liu Y, Granberg KJ, Hu L, Haapasalo H, Annala MJ, et al. IGFBP2 potentiates nuclear EGFR-STAT3 signaling. Oncogene 2016;35:738–747. ArticlePubMedPDF
- 25. Allen RE, Boxhorn LK. Regulation of skeletal muscle satellite cell proliferation and differentiation by transforming growth factor-beta, insulin-like growth factor I, and fibroblast growth factor. J Cell Physiol 1989;138:311–315. ArticlePubMed
- 26. Lynch GS, Cuffe SA, Plant DR, Gregorevic P. IGF-I treatment improves the functional properties of fast- and slow-twitch skeletal muscles from dystrophic mice. Neuromuscul Disord 2001;11:260–268. ArticlePubMed
- 27. Schertzer JD, van der Poel C, Shavlakadze T, Grounds MD, Lynch GS. Muscle-specific overexpression of IGF-I improves E-C coupling in skeletal muscle fibers from dystrophic mdx mice. Am J Physiol Cell Physiol 2008;294:C161–C168. ArticlePubMed
- 28. Aguirre GA, De Ita JR, de la Garza RG, Castilla-Cortazar I. Insulin-like growth factor-1 deficiency and metabolic syndrome. J Transl Med 2016;14:3ArticlePubMedPMC
- 29. Renehan AG, Frystyk J, Flyvbjerg A. Obesity and cancer risk: the role of the insulin-IGF axis. Trends Endocrinol Metab 2006;17:328–336. ArticlePubMed
- 30. Heald AH, Kaushal K, Siddals KW, Rudenski AS, Anderson SG, Gibson JM. Insulin-like growth factor binding protein-2 (IGFBP-2) is a marker for the metabolic syndrome. Exp Clin Endocrinol Diabetes 2006;114:371–376. ArticlePubMed
- 31. Levi J, Huynh FK, Denroche HC, Neumann UH, Glavas MM, Covey SD, et al. Hepatic leptin signalling and subdiaphragmatic vagal efferents are not required for leptin-induced increases of plasma IGF binding protein-2 (IGFBP-2) in ob/ob mice. Diabetologia 2012;55:752–762. ArticlePubMedPDF
References
Fig. 1

Insulin-like growth factor binding protein 2 (IGFBP-2) expression profile from Genecards database (www.genecards.org).
Fig. 2
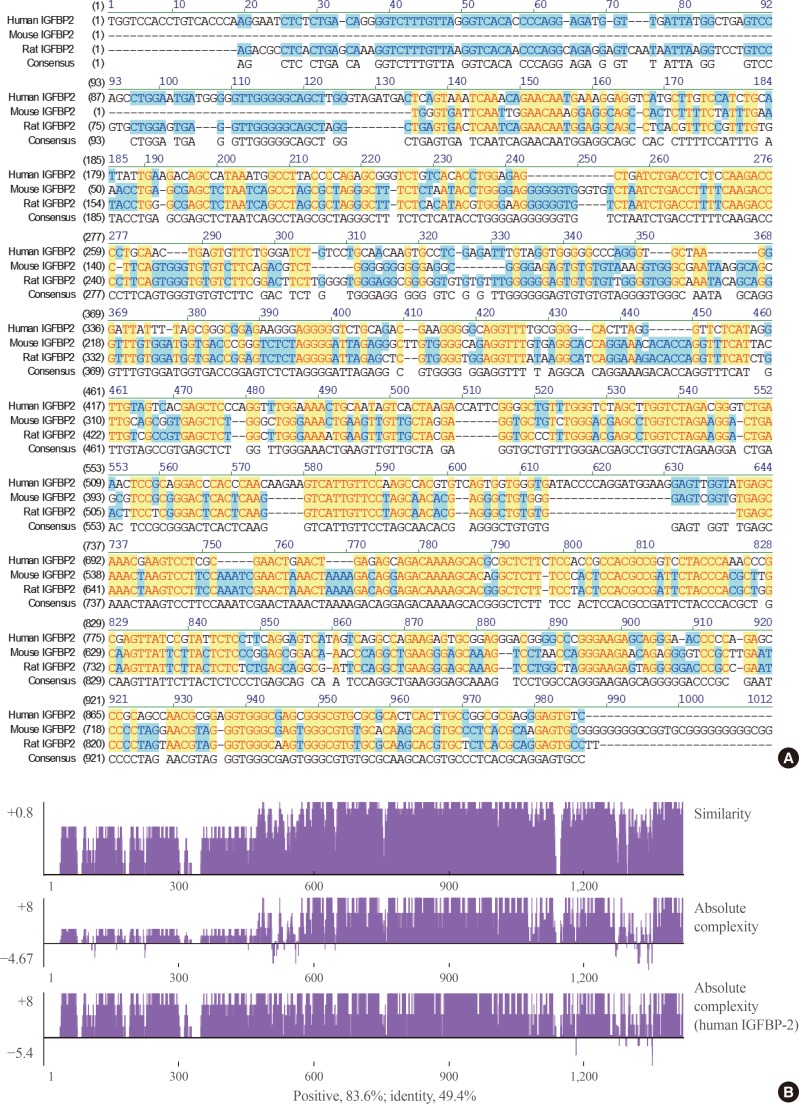
Homology comparison of the insulin-like growth factor binding protein 2 (IGFBP-2) promoter region between species which show highly conserved regions with yellow color. (A) Alignment of human, mouse and rat IGFBP-2 promoter sequences between −1,000 and 1 bp from transcription start site. (B) Similarity plots of aligned IGFBP-2 promoter sequences between species.
Fig. 3

Comparison of insulin-like growth factor binding protein 2 (IGFBP-2) exon regions between species which represent highly conserved regions with yellow color. (A) Alignment of human, mouse and rat IGFBP-2 exon sequences. (B) Similarity plots of aligned IGFBP-2 amino acids sequences between species.
Fig. 4
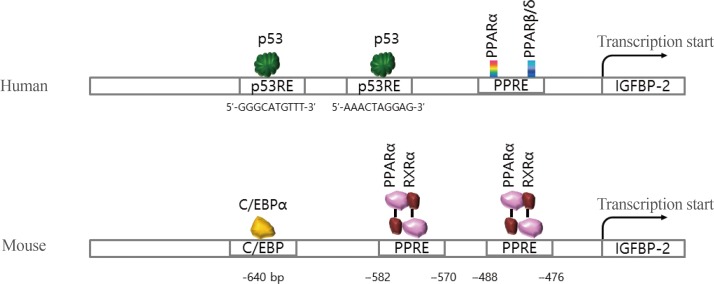
Summary of transaction factor binding to the promoter regions of the insulin-like growth factor binding protein 2 (IGFBP-2) gene for human and mouse. PPAR, peroxisome proliferation-activator receptor; PPRE, PPAR response element; C/EBPα, CCAAT-enhancer-binding protein α; RXRα, retinoid X receptor α.
Figure & Data
References
Citations
Citations to this article as recorded by 
- Predictive potential of various plasma inflammation-, angiogenesis-, and extracellular matrix remodeling-associated mediators for intra-amniotic inflammation and/or microbial invasion of the amniotic cavity in preterm labor
Eunwook Joo, Subeen Hong, Kyo Hoon Park, Hyeon Ji Kim, Min Jung Lee, Sue Shin
Archives of Gynecology and Obstetrics.2024;[Epub] CrossRef - The Role of Insulin-like Growth Factor (IGF) System in the Corneal Epithelium Homeostasis—From Limbal Epithelial Stem Cells to Therapeutic Applications
Małgorzata Woronkowicz, Harry Roberts, Piotr Skopiński
Biology.2024; 13(3): 144. CrossRef - Loss of IGFBP2 mediates alveolar type 2 cell senescence and promotes lung fibrosis
Chiahsuan Chin, Ranjithkumar Ravichandran, Kristina Sanborn, Timothy Fleming, Stephen B. Wheatcroft, Mark T. Kearney, Sofya Tokman, Rajat Walia, Michael A. Smith, David J. Flint, Thalachallour Mohanakumar, Ross M. Bremner, Angara Sureshbabu
Cell Reports Medicine.2023; 4(3): 100945. CrossRef - Circulating IGF-1 and IGFBP-2 may be biomarkers for risk stratification in patients with acute coronary syndrome: A prospective cohort study
Wei Wang, Ying Sun, De-Gang Mo, Tai Li, Heng-Chen Yao
Nutrition, Metabolism and Cardiovascular Diseases.2023; 33(9): 1740. CrossRef - Receptor Tyrosine Kinase: Still an Interesting Target to Inhibit the Proliferation of Vascular Smooth Muscle Cells
Yilin Xiong, Yan Wang, Tao Yang, Yunmei Luo, Shangfu Xu, Lisheng Li
American Journal of Cardiovascular Drugs.2023; 23(5): 497. CrossRef - Octopus death and dying
Z Yan Wang
Integrative And Comparative Biology.2023; 63(6): 1209. CrossRef - Efficient generation of functional pancreatic β cells from dental-derived stem cells via laminin-induced differentiation
Riham M. Aly, Hadeer A. Aglan, Ghada Nour Eldeen, Nadia S. Mahmoud, Eman H. Aboul-Ezz, Hanaa H. Ahmed
Journal of Genetic Engineering and Biotechnology.2022; 20(1): 85. CrossRef - Insulin-like growth factor signalling and its significance as a biomarker in fish and shellfish research
S. Chandhini, Bushra Trumboo, Seena Jose, Tincy Varghese, M. Rajesh, V. J. Rejish Kumar
Fish Physiology and Biochemistry.2021; 47(4): 1011. CrossRef - Nanomaterials for Chronic Kidney Disease Detection
Solmaz Maleki Dizaj, Aziz Eftekhari, Shakar Mammadova, Elham Ahmadian, Mohammadreza Ardalan, Soodabeh Davaran, Aygun Nasibova, Rovshan Khalilov, Mahbuba Valiyeva, Sevil Mehraliyeva, Ebrahim Mostafavi
Applied Sciences.2021; 11(20): 9656. CrossRef - Producción de las proteínas de unión al factor de crecimiento insulinoide durante el desarrollo de la fibrosis hepática por hepatitis C crónica
D. Rosique-Oramas, M. Martínez-Castillo, A. Raya, Z. Medina-Ávila, F. Aragón, J. Limón-Castillo, A. Hernández-Barragán, A. Santoyo, E. Montalvo-Javé, J.L. Pérez-Hernández, F. Higuera-de la Tijera, A. Torre, D. Kershenobich, G. Gutiérrez-Reyes
Revista de Gastroenterología de México.2020; 85(4): 390. CrossRef - Interacción cardiorrenal y evolución de la insuficiencia cardiaca. ¿Tiene un papel la proteína de unión del factor de crecimiento de tipo insulina 2?
Susana Ravassa, Javier Beaumont, Germán Cediel, Josep Lupón, Begoña López, Ramón Querejeta, Javier Díez, Antoni Bayés-Genís, Arantxa González
Revista Española de Cardiología.2020; 73(10): 835. CrossRef - Molecular identification of grass carp igfbp2 and the effect of glucose, insulin, and glucagon on igfbp2 mRNA expression
Guokun Yang, Wenli Zhao, Chaobin Qin, Liping Yang, Xiaolin Meng, Ronghua Lu, Xiao Yan, Xianglin Cao, Yanmin Zhang, Guoxing Nie
Fish Physiology and Biochemistry.2020; 46(4): 1469. CrossRef - Production of insulin-like growth factor-binding proteins during the development of hepatic fibrosis due to chronic hepatitis C
D. Rosique-Oramas, M. Martínez-Castillo, A. Raya, Z. Medina-Ávila, F. Aragón, J. Limón-Castillo, A. Hernández-Barragán, A. Santoyo, E. Montalvo-Javé, J.L. Pérez-Hernández, F. Higuera-de la Tijera, A. Torre, D. Kershenobich, G. Gutiérrez-Reyes
Revista de Gastroenterología de México (English Edition).2020; 85(4): 390. CrossRef - Cardiorenal interaction and heart failure outcomes. A role for insulin-like growth factor binding protein 2?
Susana Ravassa, Javier Beaumont, Germán Cediel, Josep Lupón, Begoña López, Ramón Querejeta, Javier Díez, Antoni Bayés-Genís, Arantxa González
Revista Española de Cardiología (English Edition).2020;[Epub] CrossRef - Integration of single-cell datasets reveals novel transcriptomic signatures of β-cells in human type 2 diabetes
Emanuele Bosi, Lorella Marselli, Carmela De Luca, Mara Suleiman, Marta Tesi, Mark Ibberson, Decio L Eizirik, Miriam Cnop, Piero Marchetti
NAR Genomics and Bioinformatics.2020;[Epub] CrossRef - Transcriptional Analysis of FOXO1, C/EBP- and PPAR-2 Genes and Their association with Obesity-Related Insulin Resistance
Boughanem, Cabrera-Mulero, Millán-Gómez, Garrido-Sánchez, Cardona, Tinahones, Moreno-Santos, Macías-González
Genes.2019; 10(9): 706. CrossRef - Immune biomarkers in maternal plasma to identify histologic chorioamnionitis in women with preterm labor
Jeong Woo Park, Kyo Hoon Park, Song Yi Kook, Young Mi Jung, Yu Mi Kim
Archives of Gynecology and Obstetrics.2019; 299(3): 725. CrossRef - Insulin like growth factor binding protein 2 (IGFBP-2) for risk prediction in patients with severe aortic stenosis undergoing Transcatheter Aortic Valve Implantation (TAVI)
Johanna Maria Muessig, Michael Lichtenauer, Bernhard Wernly, Malte Kelm, Marcus Franz, Laura Bäz, Paul Christian Schulze, Victoria Racher, Georg Zimmermann, Hans-Reiner Figulla, Vera Paar, Uta C. Hoppe, Philippe Rouet, Alexander Lauten, Christian Jung
International Journal of Cardiology.2019; 277: 54. CrossRef - Molecular cloning and expression pattern of IGFBP-2a in black porgy (Acanthopagrus schlegelii) and evolutionary analysis of IGFBP-2s in the species of Perciformes
Xinyi Zhang, Zhiyong Zhang, Zhenpeng Yu, Jiayi Li, Shuyin Chen, Ruijian Sun, Chaofeng Jia, Fei Zhu, Qian Meng, Shixia Xu
Fish Physiology and Biochemistry.2019; 45(5): 1731. CrossRef - Current Insights into the Role of the Growth Hormone-Insulin-Like Growth Factor System in Short Children Born Small for Gestational Age
Judith S. Renes, Jaap van Doorn, Anita C.S. Hokken-Koelega
Hormone Research in Paediatrics.2019; 92(1): 15. CrossRef - Insulin-Like Growth Factor (IGF) System in Liver Diseases
Agnieszka Adamek, Aldona Kasprzak
International Journal of Molecular Sciences.2018; 19(5): 1308. CrossRef - IGFBP2 is a potential biomarker in acute kidney injury (AKI) and resveratrol-loaded nanoparticles prevent AKI
Hai-Lun Li, Zhuan Yan, Zun-Ping Ke, Xiao-Feng Tian, Li-Li Zhong, Yong-Tao Lin, Yong Xu, Dong-Hui Zheng
Oncotarget.2018; 9(93): 36551. CrossRef - Multiple optic gland signaling pathways implicated in octopus maternal behaviors and death
Z. Yan Wang, Clifton W. Ragsdale
Journal of Experimental Biology.2018;[Epub] CrossRef - Insulin-Like Growth Factor-Binding Proteins of Teleost Fishes
Daniel Garcia de la Serrana, Daniel J. Macqueen
Frontiers in Endocrinology.2018;[Epub] CrossRef
 PubReader
PubReader Cite
Cite