Articles
- Page Path
- HOME > Endocrinol Metab > Volume 30(2); 2015 > Article
-
Original ArticleExendin-4 Inhibits the Expression of SEPP1 and Fetuin-A via Improvement of Palmitic Acid-Induced Endoplasmic Reticulum Stress by AMPK
- Jinmi Lee1, Seok-Woo Hong1, Se Eun Park2, Eun-Jung Rhee2, Cheol-Young Park2, Ki-Won Oh2, Sung-Woo Park2, Won-Young Lee2
-
Endocrinology and Metabolism 2015;30(2):177-184.
DOI: https://doi.org/10.3803/EnM.2015.30.2.177
Published online: June 30, 2015
1Institute of Medical Research, Kangbuk Samsung Hospital, Sungkyunkwan University School of Medicine, Seoul, Korea.
2Division of Endocrinology and Metabolism, Department of Internal Medicine, Kangbuk Samsung Hospital, Sungkyunkwan University School of Medicine, Seoul, Korea.
- Corresponding author: Won-Young Lee. Division of Endocrinology and Metabolism, Department of Internal Medicine, Kangbuk Samsung Hospital, Sungkyunkwan University School of Medicine, 29 Saemunan-ro, Jongno-gu, Seoul 110-746, Korea. Tel: +82-2-2001-2579, Fax: +82-2-2001-1588, drlwy@hanmail.net
• Received: May 8, 2014 • Revised: May 30, 2014 • Accepted: June 24, 2014
Copyright © 2015 Korean Endocrine Society
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
ABSTRACT
-
Background
- Selenoprotein P (SEPP1) and fetuin-A, both circulating liver-derived glycoproteins, are novel biomarkers for insulin resistance and nonalcoholic fatty liver disease. However, the effect of exendin-4 (Ex-4), a glucagon-like peptide-1 receptor agonist, on the expression of hepatokines, SEPP1, and fetuin-A, is unknown.
-
Methods
- The human hepatoma cell line HepG2 was treated with palmitic acid (PA; 0.4 mM) and tunicamycin (tuni; 2ug/ml) with or without exendin-4 (100 nM) for 24 hours. The change in expression of PA-induced SEPP1, fetuin-A, and endoplasmic reticulum (ER) stress markers by exendin-4 treatment were evaluated using quantitative real-time reverse transcription polymerase chain reaction and Western blotting. Transfection of cells with AMP-activated protein kinase (AMPK) small interfering RNA (siRNA) was performed to establish the effect of exendin-4-mediated AMPK in the regulation of SEPP1 and fetuin-A expression.
-
Results
- Exendin-4 reduced the expression of SEPP1, fetuin-A, and ER stress markers including PKR-like ER kinase, inositol-requiring kinase 1α, activating transcription factor 6, and C/EBP homologous protein in HepG2 cells. Exendin-4 also reduced the expression of SEPP1 and fetuin-A in cells treated with tunicamycin, an ER stress inducer. In cells treated with the AMPK activator 5-aminoidazole-4-carboxamide ribonucleotide (AICAR), the expression of hepatic SEPP1 and fetuin-A were negatively related by AMPK, which is the target of exendin-4. In addition, exendin-4 treatment did not decrease SEPP1 and fetuin-A expression in cells transfected with AMPK siRNA.
-
Conclusion
- These data suggest that exendin-4 can attenuate the expression of hepatic SEPP1 and fetuin-A via improvement of PA-induced ER stress by AMPK.
- Selenoprotein P (SeP, SEPP1) is a glycoprotein that is mainly expressed in the liver and detected in plasma. SEPP1 plays a role in the transfer of selenium from the liver to plasma and other tissues. SEPP1 transcription is stimulated by interactions among the transcription factors forkhead box protein O1 (FoxO1), hepatocyte nuclear factor 4 (HNF4) α, and peroxisome proliferator-activated receptor-γ coactivator (PGC)-1α, which stimulate hepatic gluconeogenic gene expression. On the other hand, SEPP1 is attenuated by insulin [12]. Serum and hepatic SEPP1 levels reportedly have a strong positive relationship with the development of metabolic diseases [345]. In both animal and clinical studies, SEPP1 has been shown to induce insulin resistance due to dysregulation of insulin signaling and glucose metabolism in the liver and skeletal muscle [3]. Moreover, serum SEPP1 levels are inversely related to blood adiponectin levels in patients with type 2 diabetes, whereas increased adiponectin levels have been observed in SEPP1-knockout mice [6].
- Fetuin-A (α2-Heremans-Schmid glycoprotein), a 64-kDA circulating liver-derived glycoprotein, serves as a biomarker for insulin resistance, nonalcoholic fatty liver disease (NAFLD), and cardiovascular disease [789]. Fetuin-A levels are high in human hepatocytes exposed to high levels of palmitic acid (PA), glucose, or endoplasmic reticulum (ER) stress activator thapsigargin [10]. Fetuin-A inhibits insulin-induced IRS-1 tyrosine phosphorylation, thus aggravating insulin resistance [11]. Moreover, fetuin-A-knockout mice exhibit enhanced glucose clearance, improved insulin sensitivity, and resistance to dietary fat-induced weight gain [12].
- Exendin-4 (Ex-4), a potent glucagon-like peptide-1 (GLP-1) receptor agonist, is an incretin mimetic capable of relieving insulin resistance. It can decrease plasma glucose and triglyceride levels and increase high density lipoprotein cholesterol levels. The insulin-sensitizing effect of exendin-4 has been reported in human and animal models of insulin resistance [1314]. Exendin-4 also improves metabolic syndrome via modulation of the production and release of various cytokines associated in insulin resistance, oxidative stress, apoptosis, and inflammation [15]. GLP-1 analogue reportedly increases the level of the hepatokine fibroblast growth factor 21, which is involved in insulin sensitivity and glucose and lipid homeostasis [16]. However, the effect of exendin-4 on the expression of the novel hepatokines SEPP1 and fetuin-A remains unknown. Therefore, we evaluated changes in the expression levels of SEPP1 and fetuin-A after administration of exendin-4 under ER stress.
INTRODUCTION
- Chemical reagents
- PA, tunicamycin (Tuni), exendin-4, tauroursodeoxycholic acid (TUDCA), and 5-aminoimidazole-4-carboxamide ribonucleotide (AICAR) were obtained from Sigma-Aldrich (St. Louis, MO, USA). Antibodies to fetuin-A, inositol-requiring enzyme-1α (IRE1α), CCAAT/enhancer binding homologous protein (CHOP), and β-actin were purchased from Cell Signaling Technology (Danvers, MA, USA). SEPP1 and phosphor-IRE1α (P-IRE1α) antibodies were purchased from Abcam (Cambridge, MA, USA). PKR-like endoplasmic reticulum kinase (PERK), phosphor-PERK (P-PERK), activating transcription factor 6 (ATF6), X-box binding protein 1 (XBP-1) antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
- Cell culture and transfection
- The HepG2 human hepatoma cell line (ATCC, Manassas, VA, USA) was cultured in Dulbecco's modified Eagle's medium containing 10% fetal bovine serum and 1% penicillin/streptomycin (Gibco, Grand Island, NY, USA). At approximately 85% confluence in six-well plates, cells were exposed to the ER stress inducers PA and Tuni. After 24 hours, cultured cells were treated with 100 nM exendin-4, 200 µM TUDCA, and 1 mM AICAR, after which they were incubated for 24 hours.
- For gene knockout, the cells were transfected with 10 nM small interfering RNA (siRNA) of AMP-activated protein kinase (AMPK) and scrambled control siRNA (Santa Cruz) using the lipofectamine RNAiMAX reagent according to the manufacturer's instructions (Invitrogen, Carlsbad, CA, USA). After incubation for 24 hours, the transfected cells were treated with exendin-4 (100 nM) for 24 hours.
- Total RNA isolation and real-time reverse transcription polymerase chain reaction
- Total RNA was isolated from the cells using Trizol reagent (Invitrogen) to measure the messenger RNA (mRNA) levels of the SEPP1 and fetuin-A genes. Total RNA (2 µg) was reverse-transcribed to complementary DNA using the High Capacity RNA-to-cDNA Kit (Applied Biosystems, Foster City, CA, USA). After cDNA synthesis, quantitative real-time PCR was performed using SYBR green (Roche, Lewis, UK) and specific primers (Bioneer Co., Daejeon, Korea) according to the manufacturers' instructions. To normalize the expression of the target genes, the expression of β-actin (Actb) was used as an endogenous control in the comparative Ct method (2-delta delta Ct).
- Western blot analysis
- Cells were lysed in ice-cold RIPA buffer (Cell Signaling Technology) containing protease inhibitor cocktail (Roche) and phosphatase inhibitor cocktails (Sigma-Aldrich). The lysates were incubated on ice for 30 minutes, and then centrifuged at 13,000 rpm for 20 minutes at 4℃. The protein concentration of the supernatant was quantified using the Bradford protein assay (Bio-Rad Protein Assay, BioRad, Hercules, CA, USA) with bovine serum albumin standard (Thermo Scientific, Rockford, IL, USA). Equal amounts of protein (20 µg) were electrophoresed on 4% to 12% Bis-Tris Nupage gels (Invitrogen) and transferred to polyvinylidene difluoride membranes using the iBlot Dry Blotting System (Invitrogen). After transfer, the membranes were blocked in 5% bovine serum albumin /Tris buffered saline with Tween-20 buffer for 1 hour, and then incubated overnight at 4℃ with antibodies to SEPP1, fetuin-A, IRE1α, P-IRE1α, PERK, P-PERK, ATF6, XBP-1, CHOP, and β-actin followed by incubation with horseradish peroxidase conjugated secondary antibodies. Immunoreactive bands were visualized with enhanced chemiluminescence Western blotting detection reagents (GE Healthcare, Chalfont St. Giles, UK).
- Statistical analysis
- All statistical analyses were performed using PASW Statistics version 17 (SPSS Inc., Chicago, IL, USA). The data are presented as the mean±standard error. A P value less than 0.05 was considered statistically significant.
METHODS
- Exendin-4 reduced expression of PA-increased SEPP1 and fetuin-A in human hepatocytes
- The expression of the SEPP1 and fetuin-A genes was higher in cells treated with PA alone than in the untreated controls, and significantly decreased with exendin-4 treatment in cells that did and did not undergo PA treatment (Fig. 1A). In addition, the expression of proteins exhibited a trend similar to that of their respective protein transcripts (Fig. 1B).
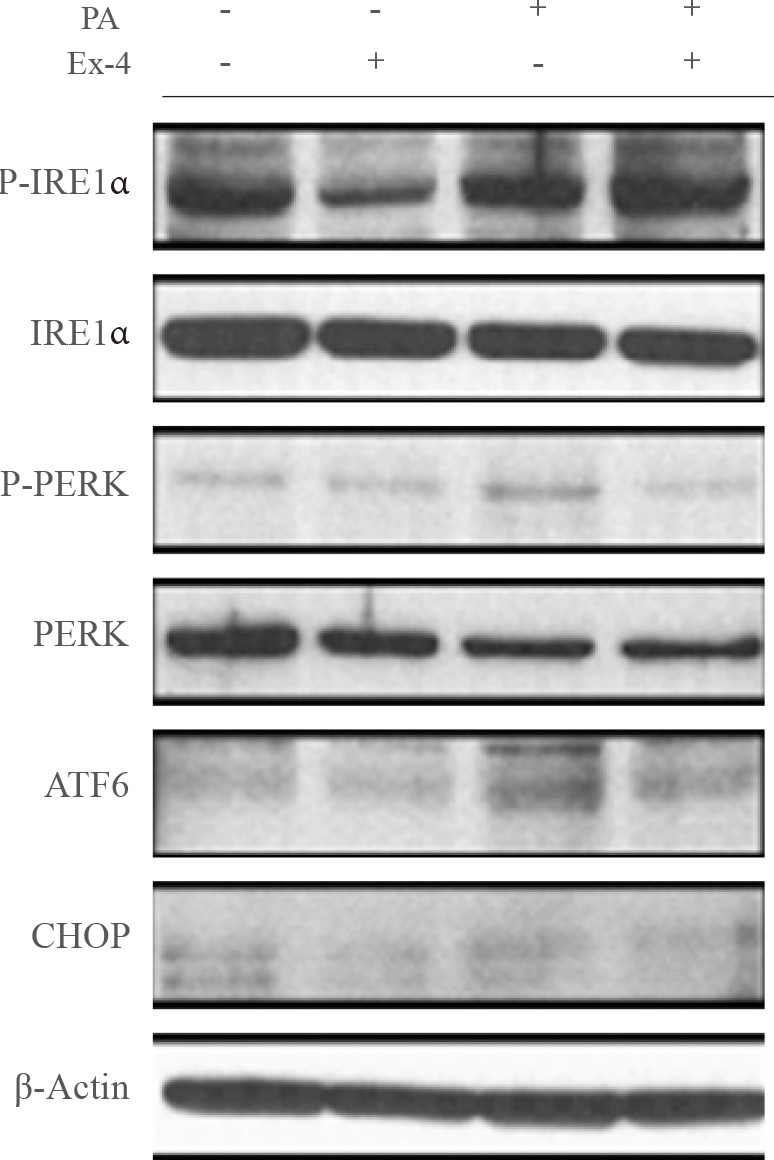
- PA, a saturated fatty acid, disrupts ER homeostasis [17], potentially leading to diabetes and hepatic steatosis. To evaluate the effect of exendin-4 on PA-induced ER stress in hepatocyte cells, we examined the effect of exendin-4 on the expression of the ER stress markers IRE1α, PERK, ATF6, and CHOP. Cells exposed to PA displayed higher P-IRE1α, P-PERK, ATF6, and CHOP protein levels, whereas PA-induced increases in the ER stress marker protein levels were reversed in cells treated with exendin-4 (Fig. 2). These results suggest that exendin-4 reduces the expression of PA-induced increases in SEPP1 and fetuin-A, and that exendin-4 improves PA-induced ER stress.
- Exendin-4 reduces expression of Tuni-induced SEPP1 and fetuin-A in human hepatocytes
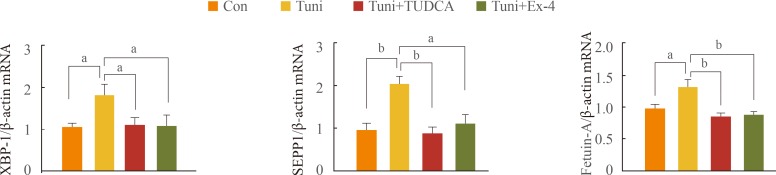
- To examine whether increased expression of SEPP1 and fetuin-A secondary to PA treatment is associated with ER stress, HepG2 cells were pretreated with Tuni, an ER stress inducer, followed by the addition of TUDCA, an ER stress inhibitor, or exendin-4. As shown in Fig. 3, the expression of SEPP1 and fetuin-A mRNA as well as XBP-1, a marker for ER stress, was significantly higher in cells treated with Tuni than in the untreated controls. In contrast, supplementation of TUDCA in cells exposed to Tuni completely abolished the effect of Tuni on the expression of these genes. Interestingly, the expression of XBP-1, SEPP1, and fetuin-A in cells treated with exendin-4 exhibited levels similar to those in cells treated with TUDCA. These data suggest that exendin-4 has a protective effect against ER stress, and that exendin-4 attenuates the expression of the SEPP1 and fetuin-A genes by relieving ER stress.
- Effect of exendin-4 on regulation of SEPP1 and fetuin-A is mediated via increased AMPK
- The AMPK activator AICAR can inhibit fatty acid-induced ER stress [18]. SIRT1-AMPK signaling induces a potent pro-tective effect of exendin-4 against fatty liver disease [19]. In this study, we have demonstrated that AICAR downregulates the expression of the SEPP1 and fetuin-A genes (Fig. 4A). We also examined whether the inhibitory effect of exendin-4 on the expression of ER stress-induced SEPP1 and fetuin-A is mediated by AMPK. The expression of AMPK increased in HepG2 cells treated with exendin-4 [19]. However, when the expression of AMPK was inhibited by AMPK siRNA, the expression of SEPP1 and fetuin-A in cells treated with exendin-4 did not decrease (Fig. 4B-D). These results suggest that exen-din-4 inhibits expression of ER stress-induced SEPP1 and fetuin-A via stimulation of AMPK. Activation of AMPK may mediate an inhibitory effect of exendin-4 on ER stress-induced hepatokines, such as SEPP1 and fetuin-A.
RESULTS
- We have demonstrated that the expression of SEPP1 and fetuin-A is significantly higher in hepatocytes treated with PA, which induces upregulation of ER stress. On the other hand, exendin-4 treatment decreases the expression of these genes via improvement of ER stress by increasing AMPK.
- Recent studies have reported that the hepatokines SEPP1 and fetuin-A can be therapeutic targets of type 2 diabetes mellitus and NAFLD [352021]. Serum SEPP1 and fetuin-A levels are reportedly associated with metabolic syndrome, which is in turn associated with hypoadiponectinemia [62122]. While patients with obesity and NAFLD show significantly increased SEPP1 and fetuin-A levels [423], SEPP1-knockout mice exhibit improved insulin sensitivity in liver and muscle and attenuated adipocyte hypertrophy [3]. Mao and Teng [24] reported that an increased plasma SEPP1 level is the result, rather than the cause of glucose dysregulation, although further studies are needed. In the present study, we demonstrated that the expression of SEPP1 and fetuin-A increased in hepatocytes treated with PA, which can induce fatty liver disease and hepatic insulin resistance [1925], and decreased in hepatocytes treated with exendin-4, which exhibits antidiabetic actions through the GLP-1 receptor. These results are consistent with those of previous studies showing that SEPP1 and fetuin-A are novel biomarkers for metabolic disorders, including obesity, diabetes, and hepatic steatosis [41026].
- ER stress plays a crucial role in obesity, insulin resistance [27], and NAFLD [28]. The ER stress response is triggered by different stimuli such as oxidative stress, hypoxia, enhanced protein synthesis, high levels of glucose and saturated fatty acids, and high levels of ER stress inducers, such as thapsigargin and Tuni [2930]. The ER stress response also stimulates ER stress sensors such as ATF6, phosphorylation of PERK and IRE-1α [31], and ER stress-induced transcription factors, such as CHOP and XBP-1 [3233]. In a previous study, we demonstrated that exendin-4 has a protective effect against PA-induced ER stress in hepatocytes [34]. In cells treated with PA, changes in the expression of the ER stress markers, SEPP1 and fetuin-A showed similar patterns, and exendin-4 treatment in cells pretreated with PA or Tuni significantly reduced the expression of these genes. In addition, relieving ER stress by exendin-4 induced decreased expression of the SEPP1 and fetuin-A genes. Thus, these results suggest that decreased expression of SEPP1 and fetuin-A in cells treated with exendin-4 may be associated with improvement of ER stress by exendin-4.
- SEPP1 expression is reportedly stimulated by the interaction between the transcription factors FoxO1 and HNF4α, and the coactivator PGC-1α, which stimulates hepatic gluconeogenic gene expression. Jung et al. [35] reported that adiponectin suppresses hepatic SEPP1 through AMPK-mediated phosphorylation of FoxO1a, and ameliorates hepatic fetuin-A through AMPK-mediated reduction of nuclear factor-κB activity [36]. Exendin-4 upregulated the expression of adiponectin and adiponectin receptor 2, and improved hepatic steatosis via SIRT1/AMPK signaling in mice models of diet-induced obesity [19]. The current study suggests that SEPP1 and fetuin-A decreased upon treatment with exendin-4 and the AMPK activator AICAR, whereas when the AMPK gene was silenced with specific siRNA, the SEPP1 and fetuin-A expression did not decrease with exendin-4 treatment. These data suggest that the effect of exendin-4 on the regulation of hepatic SEPP1 and fetuin-A may be mediated by AMPK.
- However, the regulatory mechanism of AMPK action on the expression of the SEPP1 and fetuin-A genes remains unclear. Unlike the contention by Jung et al. [35] that salsalate- and adiponectin-mediated AMPK suppresses FoxO1 activity, which is positively associated with SEPP1, Takayama et al. [37] reported that a decrease in the SEPP1 level by treatment with the AMPK activator metformin is dependent on FoxO3a, but not on FoxO1. Moreover, the mechanism of AMPK action on the regulation of fetuin-A expression is poorly defined. Further studies are needed to clarify the effect of exendin-4 on the regulation of hepatic SEPP1 and fetuin-A by treatment with AMPK.
- In conclusion, this study demonstrated that PA-induced ER stress stimulates the novel hepatokines SEPP1 and fetuin-A. This study also showed that exendin-4 can suppress the expression of hepatic SEPP1 and fetuin-A via improvement of ER stress by AMPK. Further studies are needed to clarify the details of the mechanism by which exendin-4-induced AMPK controls SEPP1 and fetuin-A.
DISCUSSION
-
Acknowledgements
- This study was supported by a National Research Foundation (NRF) grant funded by the Korean government (NRF-2013R1A 1A2063069) and by a grant from Yuhan Corporation (S-2014-1957-000).
ACKNOWLEDGMENTS
-
CONFLICTS OF INTEREST: No potential conflict of interest relevant to this article was reported.
Article information
- 1. Walter PL, Steinbrenner H, Barthel A, Klotz LO. Stimulation of selenoprotein P promoter activity in hepatoma cells by FoxO1a transcription factor. Biochem Biophys Res Commun 2008;365:316–321. ArticlePubMed
- 2. Speckmann B, Walter PL, Alili L, Reinehr R, Sies H, Klotz LO, Steinbrenner H. Selenoprotein P expression is controlled through interaction of the coactivator PGC-1alpha with FoxO1a and hepatocyte nuclear factor 4alpha transcription factors. Hepatology 2008;48:1998–2006. ArticlePubMed
- 3. Misu H, Takamura T, Takayama H, Hayashi H, Matsuzawa-Nagata N, Kurita S, Ishikura K, Ando H, Takeshita Y, Ota T, Sakurai M, Yamashita T, Mizukoshi E, Yamashita T, Honda M, Miyamoto K, Kubota T, Kubota N, Kadowaki T, Kim HJ, Lee IK, Minokoshi Y, Saito Y, Takahashi K, Yamada Y, Takakura N, Kaneko S. A liver-derived secretory protein, selenoprotein P, causes insulin resistance. Cell Metab 2010;12:483–495. ArticlePubMed
- 4. Choi HY, Hwang SY, Lee CH, Hong HC, Yang SJ, Yoo HJ, Seo JA, Kim SG, Kim NH, Baik SH, Choi DS, Choi KM. Increased selenoprotein P levels in subjects with visceral obesity and nonalcoholic fatty liver disease. Diabetes Metab J 2013;37:63–71. ArticlePubMedPMC
- 5. Yang SJ, Hwang SY, Choi HY, Yoo HJ, Seo JA, Kim SG, Kim NH, Baik SH, Choi DS, Choi KM. Serum selenoprotein P levels in patients with type 2 diabetes and prediabetes: implications for insulin resistance, inflammation, and atherosclerosis. J Clin Endocrinol Metab 2011;96:E1325–E1329. ArticlePubMed
- 6. Misu H, Ishikura K, Kurita S, Takeshita Y, Ota T, Saito Y, Takahashi K, Kaneko S, Takamura T. Inverse correlation between serum levels of selenoprotein P and adiponectin in patients with type 2 diabetes. PLoS One 2012;7:e34952ArticlePubMedPMC
- 7. Stefan N, Hennige AM, Staiger H, Machann J, Schick F, Krober SM, Machicao F, Fritsche A, Haring HU. Alpha2-Heremans-Schmid glycoprotein/fetuin-A is associated with insulin resistance and fat accumulation in the liver in humans. Diabetes Care 2006;29:853–857. ArticlePubMed
- 8. Haukeland JW, Dahl TB, Yndestad A, Gladhaug IP, Loberg EM, Haaland T, Konopski Z, Wium C, Aasheim ET, Johansen OE, Aukrust P, Halvorsen B, Birkeland KI. Fetuin A in nonalcoholic fatty liver disease: in vivo and in vitro studies. Eur J Endocrinol 2012;166:503–510. ArticlePubMed
- 9. Zhao ZW, Lin CG, Wu LZ, Luo YK, Fan L, Dong XF, Zheng H. Serum fetuin-A levels are associated with the presence and severity of coronary artery disease in patients with type 2 diabetes. Biomarkers 2013;18:160–164. ArticlePubMed
- 10. Ou HY, Wu HT, Hung HC, Yang YC, Wu JS, Chang CJ. Endoplasmic reticulum stress induces the expression of fetuin-A to develop insulin resistance. Endocrinology 2012;153:2974–2984. ArticlePubMedPDF
- 11. Srinivas PR, Wagner AS, Reddy LV, Deutsch DD, Leon MA, Goustin AS, Grunberger G. Serum alpha 2-HS-glycoprotein is an inhibitor of the human insulin receptor at the tyrosine kinase level. Mol Endocrinol 1993;7:1445–1455. ArticlePubMed
- 12. Mathews ST, Singh GP, Ranalletta M, Cintron VJ, Qiang X, Goustin AS, Jen KL, Charron MJ, Jahnen-Dechent W, Grunberger G. Improved insulin sensitivity and resistance to weight gain in mice null for the Ahsg gene. Diabetes 2002;51:2450–2458. ArticlePubMed
- 13. Ding X, Saxena NK, Lin S, Gupta NA, Anania FA. Exendin-4, a glucagon-like protein-1 (GLP-1) receptor agonist, reverses hepatic steatosis in ob/ob mice. Hepatology 2006;43:173–181. ArticlePubMed
- 14. Lam S, See S. Exenatide: a novel incretin mimetic agent for treating type 2 diabetes mellitus. Cardiol Rev 2006;14:205–211. ArticlePubMed
- 15. Dorecka M, Siemianowicz K, Francuz T, Garczorz W, Chyra A, Klych A, Romaniuk W. Exendin-4 and GLP-1 decreases induced expression of ICAM-1, VCAM-1 and RAGE in human retinal pigment epithelial cells. Pharmacol Rep 2013;65:884–890. ArticlePubMed
- 16. Yang M, Zhang L, Wang C, Liu H, Boden G, Yang G, Li L. Liraglutide increases FGF-21 activity and insulin sensitivity in high fat diet and adiponectin knockdown induced insulin resistance. PLoS One 2012;7:e48392ArticlePubMedPMC
- 17. Wei Y, Wang D, Topczewski F, Pagliassotti MJ. Saturated fatty acids induce endoplasmic reticulum stress and apoptosis independently of ceramide in liver cells. Am J Physiol Endocrinol Metab 2006;291:E275–E281. ArticlePubMed
- 18. Mayer CM, Belsham DD. Palmitate attenuates insulin signaling and induces endoplasmic reticulum stress and apoptosis in hypothalamic neurons: rescue of resistance and apoptosis through adenosine 5’ monophosphate-activated protein kinase activation. Endocrinology 2010;151:576–585. ArticlePubMedPDF
- 19. Lee J, Hong SW, Chae SW, Kim DH, Choi JH, Bae JC, Park SE, Rhee EJ, Park CY, Oh KW, Park SW, Kim SW, Lee WY. Exendin-4 improves steatohepatitis by increasing Sirt1 expression in high-fat diet-induced obese C57BL/6J mice. PLoS One 2012;7:e31394ArticlePubMedPMC
- 20. Malin SK, Del Rincon JP, Huang H, Kirwan JP. Exercise-induced lowering of fetuin-A may increase hepatic insulin sensitivity. Med Sci Sports Exerc 2014;46:2085–2090. ArticlePubMedPMC
- 21. Ishibashi A, Ikeda Y, Ohguro T, Kumon Y, Yamanaka S, Takata H, Inoue M, Suehiro T, Terada Y. Serum fetuin-A is an independent marker of insulin resistance in Japanese men. J Atheroscler Thromb 2010;17:925–933. ArticlePubMed
- 22. Hennige AM, Staiger H, Wicke C, Machicao F, Fritsche A, Haring HU, Stefan N. Fetuin-A induces cytokine expression and suppresses adiponectin production. PLoS One 2008;3:e1765ArticlePubMedPMC
- 23. Reinehr T, Roth CL. Fetuin-A and its relation to metabolic syndrome and fatty liver disease in obese children before and after weight loss. J Clin Endocrinol Metab 2008;93:4479–4485. ArticlePubMedPDF
- 24. Mao J, Teng W. The relationship between selenoprotein P and glucose metabolism in experimental studies. Nutrients 2013;5:1937–1948. ArticlePubMedPMC
- 25. Nakamura S, Takamura T, Matsuzawa-Nagata N, Takayama H, Misu H, Noda H, Nabemoto S, Kurita S, Ota T, Ando H, Miyamoto K, Kaneko S. Palmitate induces insulin resistance in H4IIEC3 hepatocytes through reactive oxygen species produced by mitochondria. J Biol Chem 2009;284:14809–14818. ArticlePubMedPMC
- 26. Jung CH, Kim BY, Kim CH, Kang SK, Jung SH, Mok JO. Associations of serum fetuin-A levels with insulin resistance and vascular complications in patients with type 2 diabetes. Diab Vasc Dis Res 2013;10:459–467. ArticlePubMed
- 27. Ozcan U, Cao Q, Yilmaz E, Lee AH, Iwakoshi NN, Ozdelen E, Tuncman G, Gorgun C, Glimcher LH, Hotamisligil GS. Endoplasmic reticulum stress links obesity, insulin action, and type 2 diabetes. Science 2004;306:457–461. ArticlePubMed
- 28. Liu J, Jin X, Yu CH, Chen SH, Li WP, Li YM. Endoplasmic reticulum stress involved in the course of lipogenesis in fatty acids-induced hepatic steatosis. J Gastroenterol Hepatol 2010;25:613–618. ArticlePubMed
- 29. Alhusaini S, McGee K, Schisano B, Harte A, McTernan P, Kumar S, Tripathi G. Lipopolysaccharide, high glucose and saturated fatty acids induce endoplasmic reticulum stress in cultured primary human adipocytes: salicylate alleviates this stress. Biochem Biophys Res Commun 2010;397:472–478. ArticlePubMed
- 30. Minamino T, Komuro I, Kitakaze M. Endoplasmic reticulum stress as a therapeutic target in cardiovascular disease. Circ Res 2010;107:1071–1082. ArticlePubMed
- 31. Schroder M, Kaufman RJ. The mammalian unfolded protein response. Annu Rev Biochem 2005;74:739–789. ArticlePubMed
- 32. Yoshida H, Matsui T, Yamamoto A, Okada T, Mori K. XBP1 mRNA is induced by ATF6 and spliced by IRE1 in response to ER stress to produce a highly active transcription factor. Cell 2001;107:881–891. ArticlePubMed
- 33. Fawcett TW, Martindale JL, Guyton KZ, Hai T, Holbrook NJ. Complexes containing activating transcription factor (ATF)/cAMP-responsive-element-binding protein (CREB) interact with the CCAAT/enhancer-binding protein (C/EBP)-ATF composite site to regulate Gadd153 expression during the stress response. Biochem J 1999;339(Pt 1):135–141. ArticlePubMedPMC
- 34. Lee J, Hong SW, Park SE, Rhee EJ, Park CY, Oh KW, Park SW, Lee WY. Exendin-4 attenuates endoplasmic reticulum stress through a SIRT1-dependent mechanism. Cell Stress Chaperones 2014;19:649–656. ArticlePubMedPMCPDF
- 35. Jung TW, Choi HY, Lee SY, Hong HC, Yang SJ, Yoo HJ, Youn BS, Baik SH, Choi KM. Salsalate and adiponectin improve palmitate-induced insulin resistance via inhibition of selenoprotein P through the AMPK-FOXO1α pathway. PLoS One 2013;8:e66529ArticlePubMedPMC
- 36. Jung TW, Youn BS, Choi HY, Lee SY, Hong HC, Yang SJ, Yoo HJ, Kim BH, Baik SH, Choi KM. Salsalate and adiponectin ameliorate hepatic steatosis by inhibition of the hepatokine fetuin-A. Biochem Pharmacol 2013;86:960–969. ArticlePubMed
- 37. Takayama H, Misu H, Iwama H, Chikamoto K, Saito Y, Murao K, Teraguchi A, Lan F, Kikuchi A, Saito R, Tajima N, Shirasaki T, Matsugo S, Miyamoto K, Kaneko S, Takamura T. Metformin suppresses expression of the selenoprotein P gene via an AMP-activated kinase (AMPK)/FoxO3a pathway in H4IIEC3 hepatocytes. J Biol Chem 2014;289:335–345. ArticlePubMed
References
Fig. 1

Exendin-4 (Ex-4) reduced the expression of selenoprotein P (SEPP1) and fetuin-A in HepG2 cells treated with palmitic acid (PA). HepG2 cells were incubated in the presence or absence of PA-containing medium, and treated with or without 100 nM Ex-4 for 24 hours. (A, B) The expression of SEPP1 and fetuin-A was analyzed using quantitative real-time reverse transcription polymerase chain reaction and Western blotting, and the data were normalized based on the β-actin. Con, control; mRNA, messenger RNA. aP<0.05; bP<0.01.
Fig. 2

Exendin-4 (Ex-4) reduced the expression of palmitic acid (PA)-induced endoplasmic reticulum stress markers. HepG2 cells were incubated in the presence or absence of PA-containing medium, and treated with or without 100 nM exendin-4 for 24 hours. Protein expression of inositol-requiring enzyme-1α (IRE1α), PKR-like endoplasmic reticulum kinase (PERK), activating transcription factor 6 (ATF6), and CCAAT/enhancer binding homologous protein (CHOP) were analyzed by Western blotting. P-IRE1α, phosphor-IRE1α; P-PERK, phosphor-PERK.
Fig. 3

Expression of selenoprotein P (SEPP1) and fetuin-A increased by endoplasmic reticulum (ER) stress was reversed by exendin-4 (Ex-4). HepG2 cells were treated with tunicamycin (Tuni), an ER stress inducer, for 24 hours, after which tauroursodeoxycholic acid (TUDCA), an ER stress inhibitor, or Ex-4 was added for 24 hours. The gene expression levels of X-box binding protein 1 (XBP-1), SEPP1, and fetuin-A were analyzed using quantitative real-time reverse transcription polymerase chain reaction, and the data were normalized based on the β-actin. Con, control; mRNA, messenger RNA. aP<0.05; bP<0.01.
Fig. 4
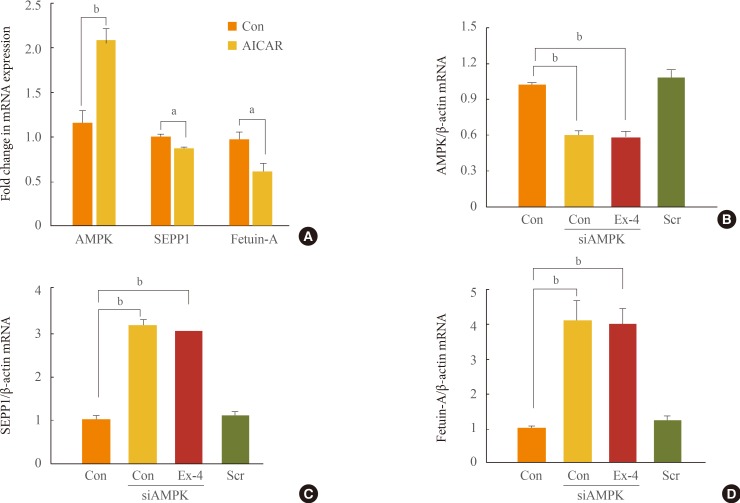
Expression of selenoprotein P (SEPP1) and fetuin-A in cells treated with exendin-4 (Ex-4) was regulated by AMP-activated protein kinase (AMPK). (A) HepG2 cells were treated with 5-aminoimidazole-4-carboxamide ribonucleotide (AICAR), an AMPK activator, for 24 hours. (B-D) Cells were transfected with the specific small interfering RNA (siRNA) for AMPK or scrambled siRNA (Scr) for 24 hours, and then added to a container with or without 100 nM Ex-4 for 24 hours. The expression of AMPK, SEPP1, and fetuin-A messenger RNA (mRNA) was measured using quantitative real-time reverse transcription polymerase chain reaction. Con, control. aP<0.05; bP<0.01.
Figure & Data
References
Citations
Citations to this article as recorded by 
- Maternal Organic Selenium Supplementation Relieves Intestinal Endoplasmic Reticulum Stress in Piglets by Enhancing the Expression of Glutathione Peroxidase 4 and Selenoprotein S
Dajiang Ding, Daolin Mou, Heng Zhu, Xuemei Jiang, Lianqiang Che, Zhengfeng Fang, Shengyu Xu, Yan Lin, Yong Zhuo, Jian Li, Chao Huang, Yuanfeng Zou, Lixia Li, De Wu, Bin Feng
Frontiers in Nutrition.2022;[Epub] CrossRef - Alliin, capsaicin, and gingerol attenuate endoplasmic reticulum stress-induced hepatic steatosis in HepG2 cells and C57BL/6N mice
Ye-Rang Yun, Ji-Eun Lee
Journal of Functional Foods.2022; 95: 105186. CrossRef - PNPLA3 I148M is involved in the variability in anti-NAFLD response to exenatide
Yunzhi Chen, Xuemei Yan, Xiao Xu, Shuhua Yuan, Fen Xu, Hua Liang
Endocrine.2020; 70(3): 517. CrossRef - Green tea extracts reduce leukocyte cell–Derived chemotaxin 2 and selenoprotein P levels in the livers of C57BL/6J mice fed a high-fat diet
Shintaro Onishi, Hidefumi Kitazawa, Shinichi Meguro, Ichiro Tokimitsu
Bioscience, Biotechnology, and Biochemistry.2018; 82(9): 1568. CrossRef - Melatonin improves insulin resistance and hepatic steatosis through attenuation of alpha‐2‐HS‐glycoprotein
Jee‐In Heo, Dae Wui Yoon, Ji Hee Yu, Nam Hoon Kim, Hye Jin Yoo, Ji A. Seo, Sin Gon Kim, Kyung Mook Choi, Sei Hyun Baik, Dong Seop Choi, Nan Hee Kim
Journal of Pineal Research.2018;[Epub] CrossRef - Palmitic acid induces ceramide accumulation, mitochondrial protein hyperacetylation, and mitochondrial dysfunction in porcine oocytes†
Nobuhiko Itami, Koumei Shirasuna, Takehito Kuwayama, Hisataka Iwata
Biology of Reproduction.2018; 98(5): 644. CrossRef - Astragaloside IV attenuates free fatty acid-induced ER stress and lipid accumulation in hepatocytes via AMPK activation
Bing Zhou, Dan-li Zhou, Xiao-hong Wei, Rong-yu Zhong, Jie Xu, Liao Sun
Acta Pharmacologica Sinica.2017; 38(7): 998. CrossRef - New Potential Targets of Glucagon-Like Peptide 1 Receptor Agonists in Pancreatic β-Cells and Hepatocytes
Won-Young Lee
Endocrinology and Metabolism.2017; 32(1): 1. CrossRef - Selenoprotein P neutralizes lipopolysaccharide and participates in hepatic cell endoplasmic reticulum stress response
Yongzhong Zhao, Shuvojit Banerjee, Ping Huang, Xinning Wang, Candece L. Gladson, Warren D. Heston, Charles B. Foster
FEBS Letters.2016; 590(24): 4519. CrossRef - Novel phenotypes of prediabetes?
Hans-Ulrich Häring
Diabetologia.2016; 59(9): 1806. CrossRef
 PubReader
PubReader Cite
Cite