Articles
- Page Path
- HOME > Endocrinol Metab > Volume 38(2); 2023 > Article
-
Review ArticleMiscellaneous Brown Adipose Tissue: Activation and Metabolism in Humans
 Keypoint
Keypoint
Brown adipose tissue is a thermogenic organ stimulated by cold and sympathetic nervous system activation. New research suggests that brown adipose tissue is active even at thermoneutrality and after meals. With its superior energy dissipation capacity compared to white adipose tissue and muscles, brown adipose tissue is being explored as a target for weight management. This review highlights human studies examining brown adipose tissue hyper-metabolism after dietary interventions and discusses nutritional agents that may recruit brown adipocytes through transdifferentiation from white adipose tissue to brown adipose tissue. -
Imane Hachemi1
 , Mueez U-Din1,2
, Mueez U-Din1,2 -
Endocrinology and Metabolism 2023;38(2):214-222.
DOI: https://doi.org/10.3803/EnM.2023.1659
Published online: March 27, 2023
1Turku PET Center, Turku University Hospital, Turku, Finland
2Turku PET Centre, University of Turku, Turku, Finland
- Corresponding author: Mueez U-Din. Turku PET Center, Turku University Hospital, University of Turku, Kiinamyllynkatu 4-6, 20520, Turku, Finland Tel: +358-41-7027723, Fax: +358-2-231-8191, E-mail: mueez.udin@utu.fi
Copyright © 2023 Korean Endocrine Society
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
- ABSTRACT
- GRAPHICAL ABSTRACT
- INTRODUCTION
- ROLE OF BROWN ADIPOSE TISSUE IN NON-SHIVERING THERMOGENESIS
- ANATOMICAL DISTRIBUTION OF BROWN ADIPOSE TISSUE
- BROWN ADIPOSE TISSUE AND DIET-INDUCED THERMOGENESIS
- ASSESSMENT OF THE PRESENCE OF BROWN ADIPOSE TISSUE WITH [18F]FDG PET
- ACTIVATION AND HYPER-METABOLISM IN BROWN ADIPOSE TISSUE
- CONCLUSIONS
- Article information
- References
ABSTRACT
- Brown adipose tissue (BAT) is a thermogenic organ contributing to non-shivering thermogenesis. BAT becomes active under cold stress via sympathetic nervous system activation. However, recent evidence has suggested that BAT may also be active at thermoneutrality and in a postprandial state. BAT has superior energy dissipation capacity compared to white adipose tissue (WAT) and muscles. Thus, it has been proposed that the recruitment and activation of additional BAT may increase the overall energy-expending capacity in humans, potentially improving current whole-body weight management strategies. Nutrition plays a central role in obesity and weight management. Thus, this review discusses human studies describing BAT hyper-metabolism after dietary interventions. Nutritional agents that can potentially recruit brown adipocytes via the process of BAT-WAT transdifferentiation are also discussed.
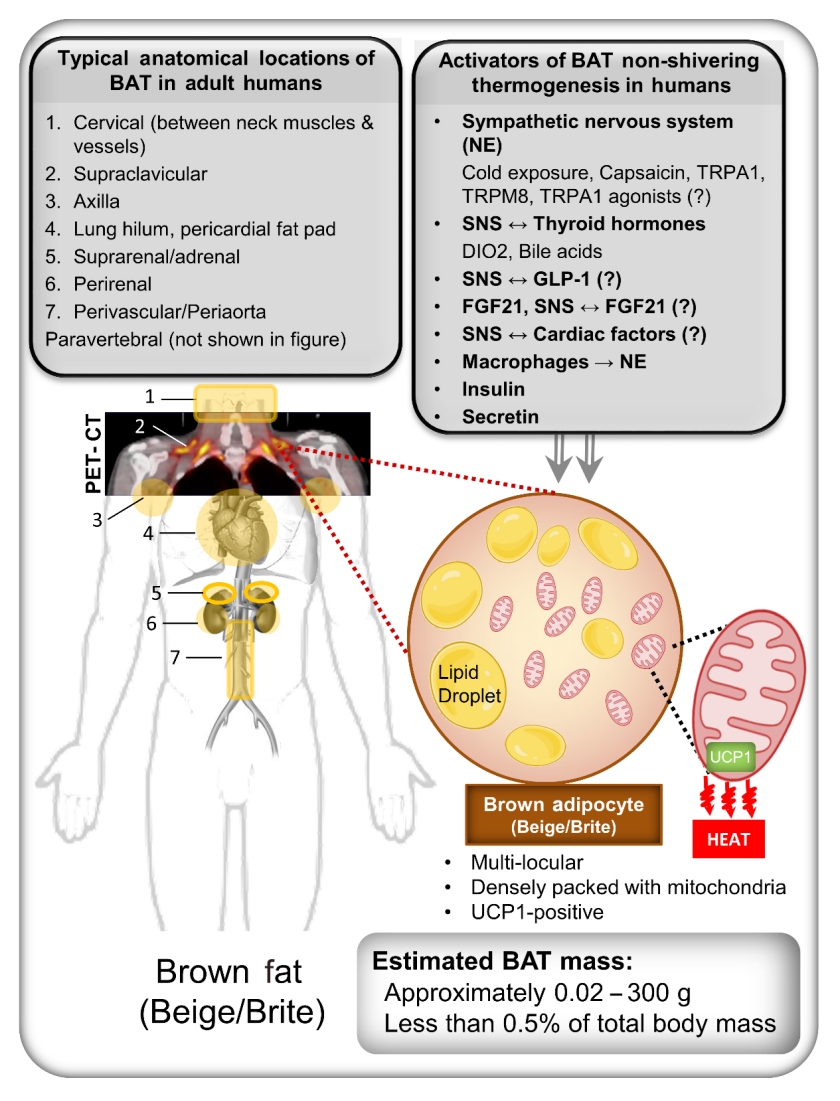
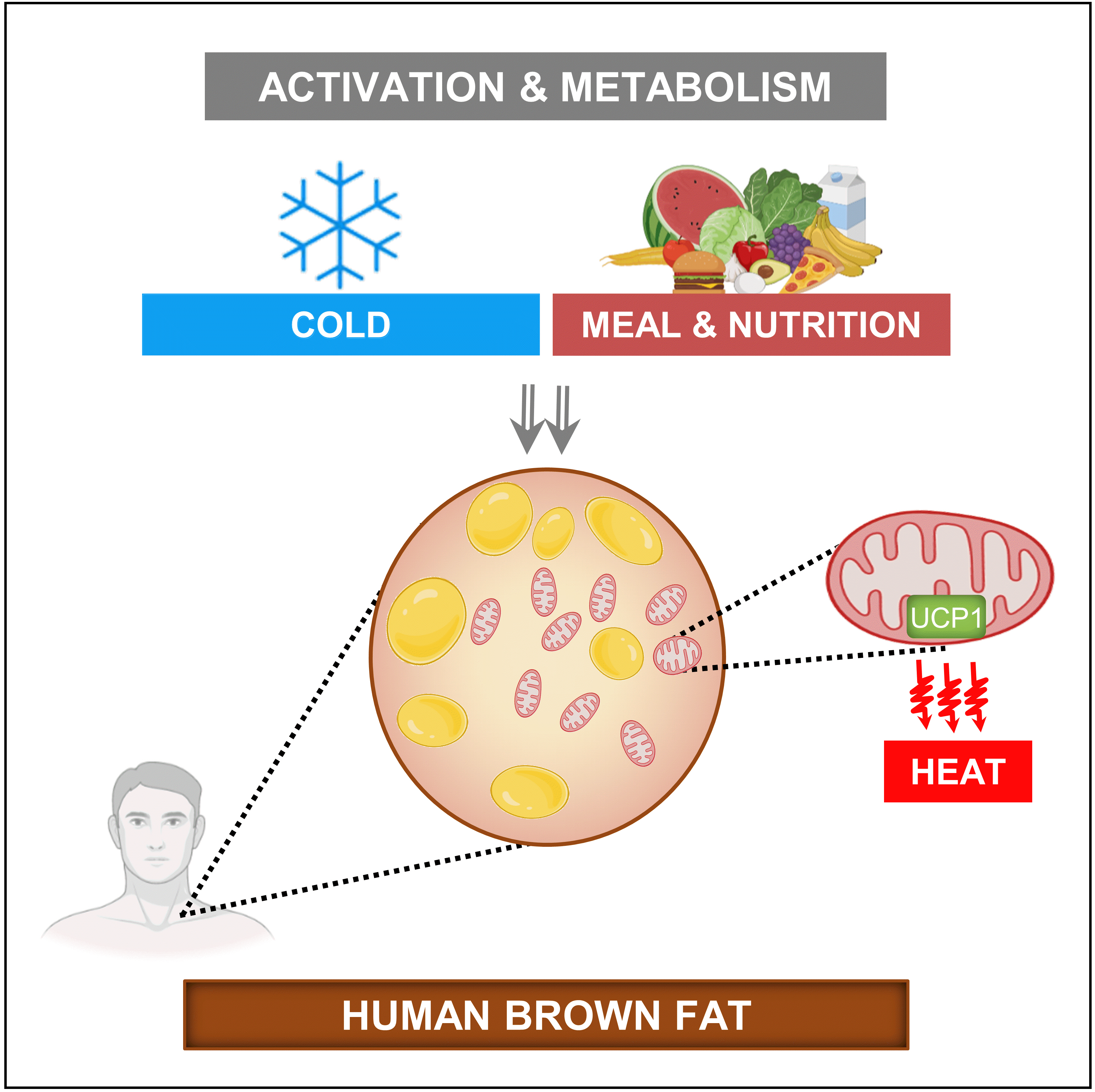
- Adipose tissue with the unique capacity to produce heat by consuming circulatory and intracellularly stored substrates is commonly known as brown adipose tissue (BAT). This adipose tissue is labelled as “brown” due to its characteristic brownish macroscopic appearance. This unique tissue morphology is due to the dissimilar cellular composition of brown adipocytes compared to white adipocytes [1]. A white adipocyte typically contains a large single lipid droplet with a few mitochondria within the cytoplasm, while a brown adipocyte characteristically possesses several small lipid droplets [2] and is densely packed with mitochondria [3]. Additionally, brown fat depots are highly vascularised [4] and innervated by abundant sympathetic nerve efferent fibres, unlike white fat depots [5,6]. The possession of multiple lipid droplets by brown adipocytes instead of a single giant lipid droplet allows the cell to increase the lipid droplet surface-to-volume ratio, which facilitates the rapid consumption of lipids in cellular respiration reactions, leading to thermogenesis [7]. The ability of brown adipocytes to produce heat arises from the presence of a large number of mitochondria, along with the high expression of uncoupling protein-1 (UCP1) [8]. Brown adipocytes express low levels of adenosine triphosphate (ATP) synthase [9], which is usually used to produce ATP by utilising the proton gradient across the mitochondrial inner membrane in the cellular respiration. In the mitochondria of brown adipocytes, the presence of high levels of UCP1, instead of ATP synthase, diminishes the proton gradient by uncoupling cellular respiration without producing ATP, and dissipating energy in the form of heat [5].
- It was previously considered that all UCP1-positive thermogenic adipocytes could be categorised as brown adipocytes. However, later evidence demonstrated that there are two types of UCP1 positive thermogenic adipocytes, which arise from distinct developmental lineages [10]. “Classical brown adipocytes” arise from engrailed-1 (En1)-expressing cells of the central dermomyotome [11] and myogenic factor 5 (Myf5)-positive progenitor cells [12], while “brown-like” adipocytes originate from Myf5-negative progenitor cells [12,13]. Classical brown adipocytes are similar to skeletal muscle in terms of their developmental origin [14] and mitochondrial proteomic signature [15], while brown-like adipocytes are closer to white adipocytes [14]. These brown-like adipocytes have been interchangeably termed as “brite” [16], “beige” [13], or “recruitable brown” [17] due to their occurrence within white adipose tissue, histologically manifesting in the form of clusters of small islands [18], and reversible white-to-brown transdifferentiation [19]. Since classical brown adipocytes and beige/brite adipocytes express unique markers of gene expressions reflecting their developmental origin [20], the evidence from genetic analysis suggests that in adult humans, brown fat depots (e.g., supraclavicular fat depots) mainly consist of beige/brite adipocytes [21], while classical brown adipocytes are usually found in rodents and human infants (e.g., interscapular fat depots) [22]. Additionally, the total amount of UCP1 protein in beige/brite adipocytes is approximately 10% of that in classical brown adipocytes; therefore, the overall UCP1-dependent thermogenic capacity of beige/brite adipocytes may be lower than that of classical brown adipocytes [23].
INTRODUCTION
- In human neonates and infants, BAT has been reported to be anatomically present in the intrascapular, supraclavicular, axillary, neck, and suprarenal areas [24]. Human neonates have a higher body surface-to-volume ratio and less muscle mass than adults; therefore, maintaining the body temperature is challenging and requires non-shivering means of heat generation. Brown fat in neonates constitutes approximately 5% of the body weight. As neonates get older, much of the BAT depots disappear, while some remain into adult life [25]. In human adults, the major sites of BAT (beige/brite) include fat depots between neck muscles; supraclavicular fat depots; depots in the axillae, the hilum of the lungs, and the area around cardiac muscles; supra- and peri-renal and adrenal depots; depots around blood vessels (e.g., the aorta) [25]; and small fat depots along the vertebral column (Fig. 1) [26]. The reported mass of brown fat in adult humans according to positron emission tomography/computed tomography (PET-CT) imaging studies is highly variable. However, the amount usually ranges from 0.02 to 300 g [27-32], which constitutes less than 0.5% of the total human body mass (75 kg). However, estimations of brown fat mass in adult humans with PET-CT are limited by the low sensitivity and spatial resolution of PET and the non-availability of brown fat-specific PET radiotracers. The traditionally used 18-F-fluorodeoxyglucose ([18F]FDG) radiotracer merely highlights the areas in the body with comparatively higher glucose metabolism; therefore, utilising the [18F]FDG radiotracer to estimate the amount and prevalence of BAT likely leads to a gross underestimation. Whilst a few studies have estimated BAT mass in humans utilising the fatty acid radiotracer 18-F-fluoro-6-thia-heptadecanoic acid ([18F]FTHA) [31], or multiple sequentially administered [18F] FDG, [18F]FTHA, and [11C]acetate (a marker of oxidative metabolism) radiotracers [33], the total BAT mass estimated from these approaches likewise appears to be less than 300 g.
ROLE OF BROWN ADIPOSE TISSUE IN NON-SHIVERING THERMOGENESIS
- The eccentric distribution of brown fat in the adult human body (Fig. 1) is likely to be of physiological importance; however, our current understanding regarding the functional relevance of human BAT location is largely speculative and/or based on rodent studies. Perivascular brown fat likely acts as an active metabolic heater for blood flowing to and from the cooler periphery [34]. Cervical BAT has been suggested to maintain the temperature of extracranial arterial blood owing to its proximity to the common carotid and vertebral arteries [26]. BAT located in the axillary and supraclavicular region likely warms the venous blood from the subclavian and jugular veins [26], and thus possibly protects against cardiac arrhythmias, since the myocardium is sensitive to lower coronary blood temperatures [35]. Likewise, the closer proximity of supraclavicular BAT to the brachial plexus, and paravertebral BAT to the spinal cord, suggests that these depots might protect against hypothermia to maintain optimal function and nerve conduction in the central and autonomic nervous system [26].
ANATOMICAL DISTRIBUTION OF BROWN ADIPOSE TISSUE
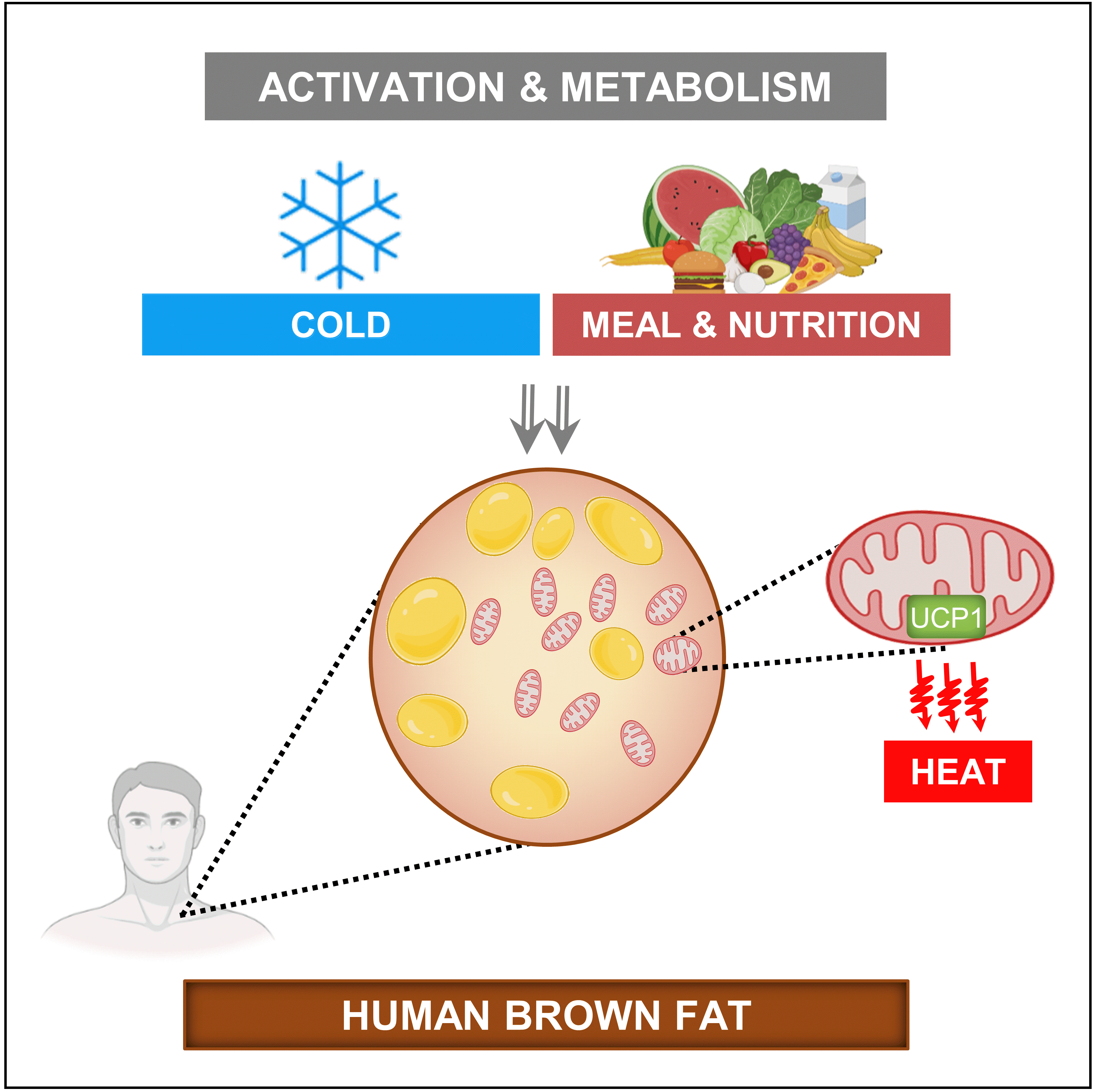
- In addition to the role of brown fat as a protector against hypothermia, brown fat tissue has been speculated to play a contributory role in diet-induced thermogenesis [36], and it has also been considered as an energy sink that helps maintain the energy balance [37]. These speculations are based on rodent studies, and the concept of “diet-induced thermogenesis” as a mechanism of maintaining energy balance in humans is controversial [38]. Nevertheless, recent evidence from a human study suggested that the extent of diet-induced thermogenesis in human supraclavicular brown fat is comparable to its cold-induced thermogenic potential [39].
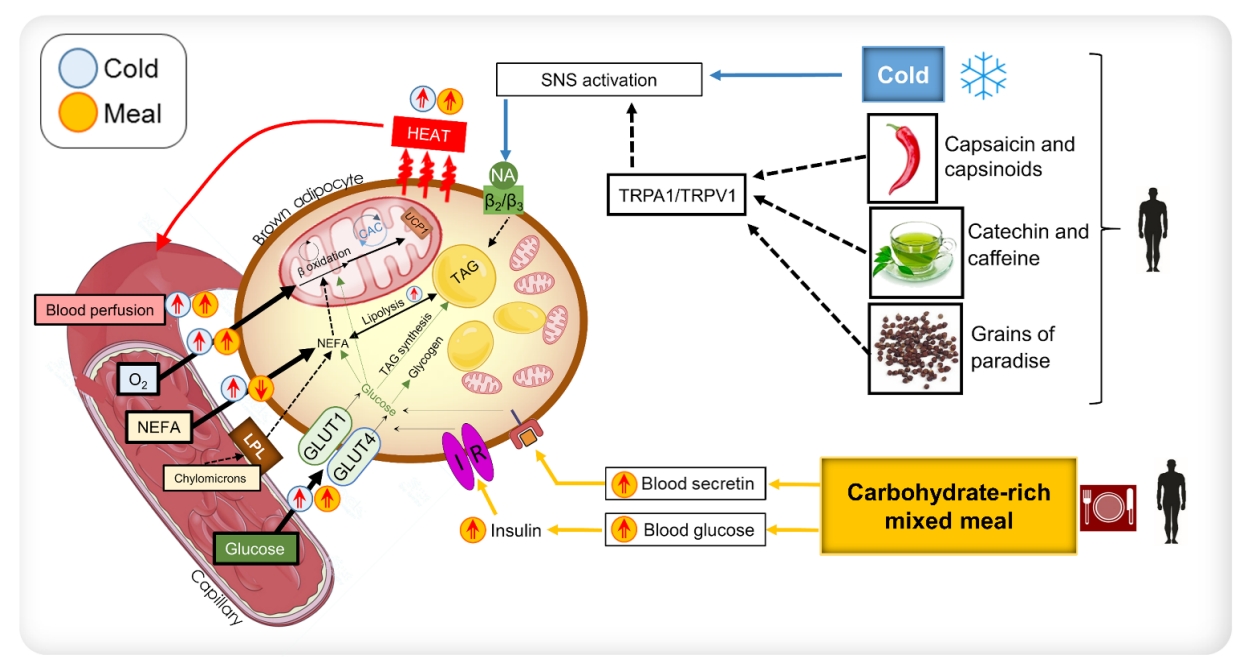
- The metabolic stimulation of brown fat for non-shivering thermogenesis in response to a cold stimulus is via a norepinephrine (NE)-dependent signal transduction pathway due to the activation of the sympathetic nervous system (SNS) [40]. However, the cold-activated increase in NE acts in a non-discriminatory fashion, exerting an effect on multiple tissues. In a quest for “selective” BAT activation, which is speculated to create a negative energy balance for weight loss without any negative effects on cardiovascular [41,42] and cognitive systems [43], a few studies have identified agents that act along with the SNS pathway or independently to stimulate BAT thermogenesis (Fig. 2) [44]. Nevertheless, studies examining the comparative contribution of brown fat in adult humans using a direct oxygen consumption PET technique have shown that supraclavicular BAT may merely contribute up to 10 to 15 kcal of energy per 100 g of available tissue when activated by a cold stimulus [31,39,45]; thus, the idea of selectively activating BAT to create a huge negative energy balance for weight loss may need to be revisited.
BROWN ADIPOSE TISSUE AND DIET-INDUCED THERMOGENESIS
- Several studies have classified individuals as “BAT-positive” or “BAT-negative” on the basis of [18F]FDG PET accumulation in the supraclavicular fat depot of humans [46-48]. These studies speculated that the differences between these two groups are largely due to the presence or absence of BAT. However, the current “gold-standard” method for assessing the presence of BAT in humans possesses some inherent limitations. The assessment of BAT glucose metabolism is merely indicative of BAT insulin sensitivity [49]; thus, the evaluation of the presence of BAT using this method and considering BAT glucose uptake as a marker of thermogenic activity has the tendency to result in over-speculative conclusions regarding the role of BAT in mealinduced thermogenesis. Dynamic PET imaging can also be used to measure the rates of glucose utilisation by BAT; however, this approach has been utilised by only a few research groups. Whole-body dynamic PET imaging with [18F]FDG has demonstrated that BAT accounts for approximately 1% of the total glucose utilisation in the body, while the skeletal muscles account for approximately 50% of the total glucose utilisation [50].
- Additionally, it has been demonstrated that a major portion of the taken-up glucose does not contribute to BAT oxidative metabolism; rather, it is used for glyceroneogenesis or de novo lipogenesis [51]. Furthermore, Weir et al. [52] showed using the microdialysis technique that in response to acute cold stimulation, the greater fraction of taken-up glucose in the human supraclavicular region is released as lactate. Hence, the use of [18F]FDG PET imaging as a marker of the presence of BAT and an indicator of thermogenic activity is not optimal.
ASSESSMENT OF THE PRESENCE OF BROWN ADIPOSE TISSUE WITH [18F]FDG PET
- The substantial uptake of glucose in the supraclavicular fat depots has been conventionally regarded as activation of BAT [53]. Hypermetabolism in BAT occurs in response to hormonal stimulation. For example, during cold stress, the activation of the SNS results in the release of NE, which acts on BAT via G protein-coupled β3-adrenergic receptors [40]. This results in a cascade of events [54] that increase mitochondrial respiration in these adipocytes. In addition to NE, other agents can stimulate brown adipocytes to enhance the thermogenic oxidative metabolism. These agents act along the NE-induced signal transduction pathway or independently. The gut hormone secretin, secreted in response to the ingestion of a meal, has been shown to stimulate BAT glucose uptake and thermogenesis via the presence of secretin receptors in human brown adipose tissue [55,56]. Furthermore, humans with hyperthyroidism also manifest hyper-metabolism of BAT [57]; however, the precise mechanism underlying this phenomenon still needs to be elucidated. Insulin has been shown to influence BAT metabolism [58,59]. Orava et al. [28] reported that BAT exhibited increased glucose uptake under conditions of a hyperinsulinemic euglycemic clamp.
- Additionally, Vosselman et al. [60] found that BAT had comparable glucose uptake after the ingestion of a high-caloric carbohydrate-rich meal. Despite this evidence, it is not known in humans whether insulin stimulation leads to increased respiratory activity in BAT or merely increased glucose uptake. The data published by U Din et al. [39] showed that oxidative metabolism in BAT increased after the consumption of a carbohydrate-dominant mixed meal, with a magnitude equivalent to that observed in cold stress. The ingestion of a carbohydrate-rich meal raises blood glucose levels, which consequently stimulates insulin production. Insulin has been suggested to enhance the expression of glucose transporter 4 (GLUT4) in BAT [61]; thus, GLUT4 stimulates glucose uptake into BAT [58,62,63]. Nonetheless, it is unclear whether insulin has a direct effect on stimulating BAT mitochondrial respiration. Insulin may induce increased thermogenesis in BAT by inhibiting the warm-sensitive neurons in the hypothalamus [64,65].
- The increased thermogenic metabolism via food nutrients acts along these pathways to stimulate BAT. Human studies have shown that certain food agents can stimulate BAT along the SNS or insulin-stimulatory pathway to enhance BAT metabolism. The diagrammatic representation in Fig. 2 shows these mechanisms of BAT activation demonstrated in human studies. This review will thus discuss these nutrients briefly.
- Capsaicin and capsinoids
- Capsaicin is the pungent compound present in chilli peppers, while capsinoids are capsaicin-like compounds present in non-pungent chilli peppers. Both capsaicin and capsinoids have been documented to increase catecholamine secretion via the activation of the SNS, consequently increasing whole-body energy expenditure in humans. Several reports have stated that the oral administration of capsaicin and capsinoids in humans can enhance whole-body energy expenditure in subjects with detectable BAT. In the study by Yoneshiro et al. [66], subjects with undetectable BAT, based on [18F]FDG standardised uptake value (SUVs), did not show increased whole-body energy expenditure after the oral ingestion of capsinoids. This finding was confirmed by Sun et al. [67]; however, the acute activation of BAT after capsinoid ingestion was not observed. Capsaicin has been documented to activate transient receptor potential cation channel subfamily V member 1 (TRPV1), thereby increasing sympathetic nerve activity. The increase in SNS activity increases the whole-body energy expenditure and fat oxidation, comparable to the response in cold conditions.
- Repeated cold stimulation in humans, or cold acclimation, has been reported to increase BAT activity and cold-induced thermogenesis (CIT) [68-71]. Similarly, Yoneshiro et al. [72] showed that repeated daily ingestion of capsinoids for 6 weeks increased CIT. However, a direct effect of the repeated ingestion of capsinoids on BAT activity based on PET imaging was not shown. Although CIT has been speculated to represent BAT activity, more recent reports have found that CIT is in fact due to the contribution of both BAT and deep muscles [31]; thus, it can be reasonably speculated that repeated BAT activation may have the capacity to improve overall metabolic health in terms of energy expenditure and the reduction of stored fat due to the release on brown adipokines (BATokines), in a similar fashion to how the repeated stimulation of muscles (e.g., strength training) releases myokines [73].
- Catechin and caffeine
- Catechin is a bioactive compound found in tea, and the health-promoting effects associated with the consumption of tea are attributed to catechin [74]. Human studies have revealed that catechin consumption demonstrates several health benefits, including increased whole-body energy expenditure, fat oxidation [75], elevated non-shivering thermogenesis potential [76] and reduction in body fat [77]. A few human studies have also evaluated the effect of long-term catechin ingestion in relation to BAT metabolism. Yoneshiro et al. [48] found that humans with detectable BAT, using [18F]FDG, demonstrated an increase in whole-body energy expenditure shortly after the ingestion of catechin-containing beverages. This effect was not found in individuals who did not demonstrate detectable BAT on PET imaging. They also found that the chronic consumption of catechins beverages increased cold-induced thermogenic capacity. However, that study did not evaluate the chronic effects of catechin on BAT metabolism. Nirengi et al. [78] found using nearinfrared spectroscopy that the density of BAT increased after the daily ingestion of catechin-rich beverages after 12 weeks; they additionally found using MR spectroscopy that extramyocellular lipid levels decreased after this intervention.
- Grains of paradise
- Grains of paradise (Aframomum melegueta [Rosco] K. Schum.) are a type of chilli pepper found commonly in Western African countries. This plant is also known as Guinea pepper or alligator pepper. It is used as a spice and also as a remedy for stomachache, diarrhoea, and snake bite [79]. Like capsaicin, this plant is also known to activate TRPV1 channels in humans [80], thereby stimulating SNS activity [81]. Sugita et al. [47] demonstrated in humans that the oral ingestion of grains of paradise leads to an increase in whole-body energy expenditure in individuals with detectable BAT, based on [18F]FDG PET imaging.
ACTIVATION AND HYPER-METABOLISM IN BROWN ADIPOSE TISSUE
- Although BAT in adult humans can be stimulated via several nutrients found in foods, the known mechanism of BAT activation remains SNS activation and stimulation with insulin and secretin. The measurement of SUVs with [18F]FDG PET imaging does not provide a clear indication of thermogenic BAT activity; thus, [18F]FDG PET imaging findings should be extrapolated with caution. Additionally, the activation of BAT as a thermogenic agent might not be enough to overcome obesity in humans; thus, along with the activation of BAT, the influence of BAT activation on metabolism in muscle or other organs should also be simultaneously explored.
CONCLUSIONS
-
CONFLICTS OF INTEREST
No potential conflict of interest relevant to this article was reported.
-
AUTHOR CONTRIBUTIONS
The research of Mueez U-Din is financially supported by the Academy of Finland, Turku University Hospital Research Funds, Finnish Diabetes Research Foundation, Finnish Cultural Foundation, Finnish-Norwegian Medical Research Foundation, Juhani Aho Foundation for Medical Research, and Jalmari ja Rauha Ahokkaan Säätiö. The authors thank all the funding sources for supporting the research and also the colleagues who encouraged writing this review article.
Article information
- 1. Cinti S. The adipose organ. Prostaglandins Leukot Essent Fatty Acids 2005;73:9–15.ArticlePubMed
- 2. Cinti S. The role of brown adipose tissue in human obesity. Nutr Metab Cardiovasc Dis 2006;16:569–74.ArticlePubMed
- 3. Lindberg O, de Pierre J, Rylander E, Afzelius BA. Studies of the mitochondrial energy-transfer system of brown adipose tissue. J Cell Biol 1967;34:293–310.ArticlePubMedPMCPDF
- 4. Hausberger FX, Widelitz MM. Distribution of labeled erythrocytes in adipose tissue and muscle in the rat. Am J Physiol 1963;204:649–52.ArticlePubMed
- 5. Cannon B, Nedergaard J. Brown adipose tissue: function and physiological significance. Physiol Rev 2004;84:277–359.ArticlePubMed
- 6. Bartness TJ, Vaughan CH, Song CK. Sympathetic and sensory innervation of brown adipose tissue. Int J Obes (Lond) 2010;34 Suppl 1:S36–42.ArticlePubMedPMCPDF
- 7. Cinti S. The adipose organ at a glance. Dis Model Mech 2012;5:588–94.ArticlePubMedPMCPDF
- 8. Lean ME, James WP. Uncoupling protein in human brown adipose tissue mitochondria: isolation and detection by specific antiserum. FEBS Lett 1983;163:235–40.PubMed
- 9. Kramarova TV, Shabalina IG, Andersson U, Westerberg R, Carlberg I, Houstek J, et al. Mitochondrial ATP synthase levels in brown adipose tissue are governed by the c-Fo subunit P1 isoform. FASEB J 2008;22:55–63.ArticlePubMedPDF
- 10. Wu J, Bostrom P, Sparks LM, Ye L, Choi JH, Giang AH, et al. Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human. Cell 2012;150:366–76.ArticlePubMedPMC
- 11. Atit R, Sgaier SK, Mohamed OA, Taketo MM, Dufort D, Joyner AL, et al. Beta-catenin activation is necessary and sufficient to specify the dorsal dermal fate in the mouse. Dev Biol 2006;296:164–76.PubMed
- 12. Seale P, Bjork B, Yang W, Kajimura S, Chin S, Kuang S, et al. PRDM16 controls a brown fat/skeletal muscle switch. Nature 2008;454:961–7.ArticlePubMedPMCPDF
- 13. Ishibashi J, Seale P. Medicine: beige can be slimming. Science 2010;328:1113–4.ArticlePubMedPMC
- 14. Timmons JA, Wennmalm K, Larsson O, Walden TB, Lassmann T, Petrovic N, et al. Myogenic gene expression signature establishes that brown and white adipocytes originate from distinct cell lineages. Proc Natl Acad Sci U S A 2007;104:4401–6.ArticlePubMedPMC
- 15. Forner F, Kumar C, Luber CA, Fromme T, Klingenspor M, Mann M. Proteome differences between brown and white fat mitochondria reveal specialized metabolic functions. Cell Metab 2009;10:324–35.ArticlePubMed
- 16. Petrovic N, Walden TB, Shabalina IG, Timmons JA, Cannon B, Nedergaard J. Chronic peroxisome proliferator-activated receptor gamma (PPARgamma) activation of epididymally derived white adipocyte cultures reveals a population of thermogenically competent, UCP1-containing adipocytes molecularly distinct from classic brown adipocytes. J Biol Chem 2010;285:7153–64.PubMed
- 17. Enerback S. The origins of brown adipose tissue. N Engl J Med 2009;360:2021–3.ArticlePubMed
- 18. Cypess AM, Kahn CR. Brown fat as a therapy for obesity and diabetes. Curr Opin Endocrinol Diabetes Obes 2010;17:143–9.ArticlePubMedPMC
- 19. Cinti S. Transdifferentiation properties of adipocytes in the adipose organ. Am J Physiol Endocrinol Metab 2009;297:E977–86.ArticlePubMed
- 20. Walden TB, Hansen IR, Timmons JA, Cannon B, Nedergaard J. Recruited vs. nonrecruited molecular signatures of brown, “brite,” and white adipose tissues. Am J Physiol Endocrinol Metab 2012;302:E19–31.ArticlePubMed
- 21. Sharp LZ, Shinoda K, Ohno H, Scheel DW, Tomoda E, Ruiz L, et al. Human BAT possesses molecular signatures that resemble beige/brite cells. PLoS One 2012;7:e49452.ArticlePubMedPMC
- 22. Lidell ME, Betz MJ, Dahlqvist Leinhard O, Heglind M, Elander L, Slawik M, et al. Evidence for two types of brown adipose tissue in humans. Nat Med 2013;19:631–4.ArticlePubMedPDF
- 23. Nedergaard J, Cannon B. UCP1 mRNA does not produce heat. Biochim Biophys Acta 2013;1831:943–9.ArticlePubMed
- 24. Aherne W, Hull D. Brown adipose tissue and heat production in the newborn infant. J Pathol Bacteriol 1966;91:223–34.ArticlePubMed
- 25. Heaton JM. The distribution of brown adipose tissue in the human. J Anat 1972;112(Pt 1):35–9.PubMedPMC
- 26. Sacks H, Symonds ME. Anatomical locations of human brown adipose tissue: functional relevance and implications in obesity and type 2 diabetes. Diabetes 2013;62:1783–90.PubMedPMC
- 27. Cypess AM, Lehman S, Williams G, Tal I, Rodman D, Goldfine AB, et al. Identification and importance of brown adipose tissue in adult humans. N Engl J Med 2009;360:1509–17.ArticlePubMedPMC
- 28. Orava J, Nuutila P, Lidell ME, Oikonen V, Noponen T, Viljanen T, et al. Different metabolic responses of human brown adipose tissue to activation by cold and insulin. Cell Metab 2011;14:272–9.ArticlePubMed
- 29. van Marken Lichtenbelt WD, Vanhommerig JW, Smulders NM, Drossaerts JM, Kemerink GJ, Bouvy ND, et al. Coldactivated brown adipose tissue in healthy men. N Engl J Med 2009;360:1500–8.ArticlePubMed
- 30. Muzik O, Mangner TJ, Leonard WR, Kumar A, Janisse J, Granneman JG. 15O PET measurement of blood flow and oxygen consumption in cold-activated human brown fat. J Nucl Med 2013;54:523–31.ArticlePubMed
- 31. U Din M, Raiko J, Saari T, Kudomi N, Tolvanen T, Oikonen V, et al. Human brown adipose tissue [15O]O2 PET imaging in the presence and absence of cold stimulus. Eur J Nucl Med Mol Imaging 2016;43:1878–86.ArticlePubMedPMCPDF
- 32. GerngroB C, Schretter J, Klingenspor M, Schwaiger M, Fromme T. Active brown fat during 18F-FDG PET/CT imaging defines a patient group with characteristic traits and an increased probability of brown fat redetection. J Nucl Med 2017;58:1104–10.ArticlePubMed
- 33. Ouellet V, Labbe SM, Blondin DP, Phoenix S, Guerin B, Haman F, et al. Brown adipose tissue oxidative metabolism contributes to energy expenditure during acute cold exposure in humans. J Clin Invest 2012;122:545–52.ArticlePubMedPMC
- 34. Smith RE. Thermoregulatory and adaptive behavior of brown adipose tissue. Science 1964;146:1686–9.ArticlePubMed
- 35. Mattu A, Brady WJ, Perron AD. Electrocardiographic manifestations of hypothermia. Am J Emerg Med 2002;20:314–26.ArticlePubMed
- 36. Glick Z, Teague RJ, Bray GA. Brown adipose tissue: thermic response increased by a single low protein, high carbohydrate meal. Science 1981;213:1125–7.ArticlePubMed
- 37. Rothwell NJ, Stock MJ. A role for brown adipose tissue in diet-induced thermogenesis. Nature 1979;281:31–5.ArticlePubMedPDF
- 38. Kozak LP. Brown fat and the myth of diet-induced thermogenesis. Cell Metab 2010;11:263–7.ArticlePubMedPMC
- 39. U Din M, Saari T, Raiko J, Kudomi N, Maurer SF, Lahesmaa M, et al. Postprandial oxidative metabolism of human brown fat indicates thermogenesis. Cell Metab 2018;28:207–16.ArticlePubMed
- 40. Young JB, Saville E, Rothwell NJ, Stock MJ, Landsberg L. Effect of diet and cold exposure on norepinephrine turnover in brown adipose tissue of the rat. J Clin Invest 1982;69:1061–71.ArticlePubMedPMC
- 41. Fares A. Winter cardiovascular diseases phenomenon. N Am J Med Sci 2013;5:266–79.ArticlePubMedPMC
- 42. Florea VG, Cohn JN. The autonomic nervous system and heart failure. Circ Res 2014;114:1815–26.ArticlePubMed
- 43. Moreira FA, Crippa JA. The psychiatric side-effects of rimonabant. Braz J Psychiatry 2009;31:145–53.ArticlePubMed
- 44. Villarroya F, Vidal-Puig A. Beyond the sympathetic tone: the new brown fat activators. Cell Metab 2013;17:638–43.ArticlePubMed
- 45. Muzik O, Mangner TJ, Granneman JG. Assessment of oxidative metabolism in brown fat using PET imaging. Front Endocrinol (Lausanne) 2012;3:15.ArticlePubMedPMC
- 46. Hibi M, Oishi S, Matsushita M, Yoneshiro T, Yamaguchi T, Usui C, et al. Brown adipose tissue is involved in diet-induced thermogenesis and whole-body fat utilization in healthy humans. Int J Obes (Lond) 2016;40:1655–61.ArticlePubMedPMCPDF
- 47. Sugita J, Yoneshiro T, Hatano T, Aita S, Ikemoto T, Uchiwa H, et al. Grains of paradise (Aframomum melegueta) extract activates brown adipose tissue and increases whole-body energy expenditure in men. Br J Nutr 2013;110:733–8.ArticlePubMed
- 48. Yoneshiro T, Matsushita M, Hibi M, Tone H, Takeshita M, Yasunaga K, et al. Tea catechin and caffeine activate brown adipose tissue and increase cold-induced thermogenic capacity in humans. Am J Clin Nutr 2017;105:873–81.ArticlePubMed
- 49. Carpentier AC, Blondin DP, Virtanen KA, Richard D, Haman F, Turcotte EE. Brown adipose tissue energy metabolism in humans. Front Endocrinol (Lausanne) 2018;9:447.ArticlePubMedPMC
- 50. Blondin DP, Labbe SM, Phoenix S, Guerin B, Turcotte EE, Richard D, et al. Contributions of white and brown adipose tissues and skeletal muscles to acute cold-induced metabolic responses in healthy men. J Physiol 2015;593:701–14.ArticlePubMedPMC
- 51. Ma SW, Foster DO. Uptake of glucose and release of fatty acids and glycerol by rat brown adipose tissue in vivo. Can J Physiol Pharmacol 1986;64:609–14.ArticlePubMed
- 52. Weir G, Ramage LE, Akyol M, Rhodes JK, Kyle CJ, Fletcher AM, et al. Substantial metabolic activity of human brown adipose tissue during warm conditions and cold-induced lipolysis of local triglycerides. Cell Metab 2018;27:1348–55.ArticlePubMedPMC
- 53. Cypess AM, Weiner LS, Roberts-Toler C, Franquet Elia E, Kessler SH, Kahn PA, et al. Activation of human brown adipose tissue by a β3-adrenergic receptor agonist. Cell Metab 2015;21:33–8.ArticlePubMedPMC
- 54. Townsend KL, Tseng YH. Brown fat fuel utilization and thermogenesis. Trends Endocrinol Metab 2014;25:168–77.ArticlePubMedPMC
- 55. Li Y, Schnabl K, Gabler SM, Willershauser M, Reber J, Karlas A, et al. Secretin-activated brown fat mediates prandial thermogenesis to induce satiation. Cell 2018;175:1561–74.ArticlePubMed
- 56. Laurila S, Sun L, Lahesmaa M, Schnabl K, Laitinen K, Klen R, et al. Secretin activates brown fat and induces satiation. Nat Metab 2021;3:798–809.ArticlePubMedPDF
- 57. Lahesmaa M, Orava J, Schalin-Jantti C, Soinio M, Hannukainen JC, Noponen T, et al. Hyperthyroidism increases brown fat metabolism in humans. J Clin Endocrinol Metab 2014;99:E28–35.ArticlePubMed
- 58. Ebner S, Burnol AF, Ferre P, de Saintaurin MA, Girard J. Effects of insulin and norepinephrine on glucose transport and metabolism in rat brown adipocytes: potentiation by insulin of norepinephrine-induced glucose oxidation. Eur J Biochem 1987;170:469–74.ArticlePubMed
- 59. Inokuma K, Ogura-Okamatsu Y, Toda C, Kimura K, Yamashita H, Saito M. Uncoupling protein 1 is necessary for norepinephrine-induced glucose utilization in brown adipose tissue. Diabetes 2005;54:1385–91.ArticlePubMedPDF
- 60. Vosselman MJ, Brans B, van der Lans AA, Wierts R, van Baak MA, Mottaghy FM, et al. Brown adipose tissue activity after a high-calorie meal in humans. Am J Clin Nutr 2013;98:57–64.ArticlePubMed
- 61. Teruel T, Valverde AM, Benito M, Lorenzo M. Insulin-like growth factor I and insulin induce adipogenic-related gene expression in fetal brown adipocyte primary cultures. Biochem J 1996;319(Pt 2):627–32.ArticlePubMedPMCPDF
- 62. Klein J, Fasshauer M, Klein HH, Benito M, Kahn CR. Novel adipocyte lines from brown fat: a model system for the study of differentiation, energy metabolism, and insulin action. Bioessays 2002;24:382–8.ArticlePubMed
- 63. Omatsu-Kanbe M, Zarnowski MJ, Cushman SW. Hormonal regulation of glucose transport in a brown adipose cell preparation isolated from rats that shows a large response to insulin. Biochem J 1996;315(Pt 1):25–31.ArticlePubMedPMCPDF
- 64. Labbe SM, Caron A, Lanfray D, Monge-Rofarello B, Bartness TJ, Richard D. Hypothalamic control of brown adipose tissue thermogenesis. Front Syst Neurosci 2015;9:150.ArticlePubMedPMC
- 65. Sanchez-Alavez M, Tabarean IV, Osborn O, Mitsukawa K, Schaefer J, Dubins J, et al. Insulin causes hyperthermia by direct inhibition of warm-sensitive neurons. Diabetes 2010;59:43–50.ArticlePubMedPMCPDF
- 66. Yoneshiro T, Aita S, Matsushita M, Kameya T, Nakada K, Kawai Y, et al. Brown adipose tissue, whole-body energy expenditure, and thermogenesis in healthy adult men. Obesity (Silver Spring) 2011;19:13–6.ArticlePubMedPDF
- 67. Sun L, Camps SG, Goh HJ, Govindharajulu P, Schaefferkoetter JD, Townsend DW, et al. Capsinoids activate brown adipose tissue (BAT) with increased energy expenditure associated with subthreshold 18-fluorine fluorodeoxyglucose uptake in BAT-positive humans confirmed by positron emission tomography scan. Am J Clin Nutr 2018;107:62–70.ArticlePubMed
- 68. Blondin DP, Labbe SM, Tingelstad HC, Noll C, Kunach M, Phoenix S, et al. Increased brown adipose tissue oxidative capacity in cold-acclimated humans. J Clin Endocrinol Metab 2014;99:E438–46.ArticlePubMedPMC
- 69. Hanssen MJ, Hoeks J, Brans B, van der Lans AA, Schaart G, van den Driessche JJ, et al. Short-term cold acclimation improves insulin sensitivity in patients with type 2 diabetes mellitus. Nat Med 2015;21:863–5.ArticlePubMedPDF
- 70. Hanssen MJ, van der Lans AA, Brans B, Hoeks J, Jardon KM, Schaart G, et al. Short-term cold acclimation recruits brown adipose tissue in obese humans. Diabetes 2016;65:1179–89.ArticlePubMedPDF
- 71. van der Lans AA, Hoeks J, Brans B, Vijgen GH, Visser MG, Vosselman MJ, et al. Cold acclimation recruits human brown fat and increases nonshivering thermogenesis. J Clin Invest 2013;123:3395–403.ArticlePubMedPMC
- 72. Yoneshiro T, Aita S, Matsushita M, Kayahara T, Kameya T, Kawai Y, et al. Recruited brown adipose tissue as an antiobesity agent in humans. J Clin Invest 2013;123:3404–8.ArticlePubMedPMC
- 73. So B, Kim HJ, Kim J, Song W. Exercise-induced myokines in health and metabolic diseases. Integr Med Res 2014;3:172–9.ArticlePubMedPMC
- 74. Higdon JV, Frei B. Tea catechins and polyphenols: health effects, metabolism, and antioxidant functions. Crit Rev Food Sci Nutr 2003;43:89–143.ArticlePubMed
- 75. Dulloo AG, Duret C, Rohrer D, Girardier L, Mensi N, Fathi M, et al. Efficacy of a green tea extract rich in catechin polyphenols and caffeine in increasing 24-h energy expenditure and fat oxidation in humans. Am J Clin Nutr 1999;70:1040–5.ArticlePubMed
- 76. Gosselin C, Haman F. Effects of green tea extracts on nonshivering thermogenesis during mild cold exposure in young men. Br J Nutr 2013;110:282–8.ArticlePubMed
- 77. Nagao T, Komine Y, Soga S, Meguro S, Hase T, Tanaka Y, et al. Ingestion of a tea rich in catechins leads to a reduction in body fat and malondialdehyde-modified LDL in men. Am J Clin Nutr 2005;81:122–9.ArticlePubMed
- 78. Nirengi S, Amagasa S, Homma T, Yoneshiro T, Matsumiya S, Kurosawa Y, et al. Daily ingestion of catechin-rich beverage increases brown adipose tissue density and decreases extramyocellular lipids in healthy young women. Springerplus 2016;5:1363.ArticlePubMedPMCPDF
- 79. Akendengue B, Louis AM. Medicinal plants used by the Masango people in Gabon. J Ethnopharmacol 1994;41:193–200.ArticlePubMed
- 80. Riera CE, Menozzi-Smarrito C, Affolter M, Michlig S, Munari C, Robert F, et al. Compounds from Sichuan and Melegueta peppers activate, covalently and non-covalently, TRPA1 and TRPV1 channels. Br J Pharmacol 2009;157:1398–409.ArticlePubMedPMC
- 81. Iwami M, Mahmoud FA, Shiina T, Hirayama H, Shima T, Sugita J, et al. Extract of grains of paradise and its active principle 6-paradol trigger thermogenesis of brown adipose tissue in rats. Auton Neurosci 2011;161:63–7.ArticlePubMed
References
Figure & Data
References
Citations
- Spermidine activates adipose tissue thermogenesis through autophagy and fibroblast growth factor 21
Yinhua Ni, Liujie Zheng, Liqian Zhang, Jiamin Li, Yuxiang Pan, Haimei Du, Zhaorong Wang, Zhengwei Fu
The Journal of Nutritional Biochemistry.2024; 125: 109569. CrossRef - A natural sustained-intestinal release formulation of red chili pepper extracted capsaicinoids (Capsifen®) safely modulates energy balance and endurance performance: a randomized, double-blind, placebo-controlled study
N. Roopashree, Das S. Syam, I. M. Krishnakumar, K. N. Mala, Bradley S. Fleenor, Jestin Thomas
Frontiers in Nutrition.2024;[Epub] CrossRef - MRI Methods to Visualize and Quantify Adipose Tissue in Health and Disease
Katerina Nikiforaki, Kostas Marias
Biomedicines.2023; 11(12): 3179. CrossRef
 PubReader
PubReader ePub Link
ePub Link Cite
Cite